Схема подключения электроусилителя руля приора
Продолжение истории про установку ЭУР от приоры…
В предыдущей записи я постарался подробно расписать как установить ЭУР, а теперь расскажу как его подключить, а также расскажу как делать самостоятельную диагностину. (об этом в конце)
И так, для подключения ЭУР понадобится:
1. провод сечением примерно два квадрата (ну это я так на глаз), короче не тоньше чем идут на замок зажигания, три метра (это с запасом)
2. провод сечением 0,75, 8 метров
3. мамы большие, для подключения подрулевых переключателей, 30шт(с запсом)
4. мамы+папы малые, для подключения проводов с эур и ещё пару мелочей, 20шт(с запасом)
5. держатель предохранителя + предохранитель, 1шт (говорят минимум на 50 ампер, я уже езжу неделю с 30 амперным)
6. гофра, чтобы всё было аккуратно
7.
8. плоскогубцы, кусачки
9. отвёртки + и –
10. дрель, сверло
11. напильник
Вроде всё назвал.
И так первым делом тянем силовой провод на ЭУР от АКБ. Я его протянул через резинку под лобовым стеклом.
Держатель предохранителя ставим как можно ближе к АКБ.
Сам предохранитель пока не ставим. Минусовой провод кинул на переходную пластину.
Далее прокидываем три провода до датчика скорости, +, — и сигнальный. Главное при подключении смотрите распиновку на своём датчике, там подписано, что куда идёт. Плюсовой провод от датчика я подцепил на верхний блок предохранителей, на крайний левый.
Минус взял со шпильки которая держит панель. К сигнальному прикрепил маленькую клемму и вывел к щитку приборов.
Далее подключаем остальные провода ЭУР по схеме
. К штатным проводам я припаивал клеммы, для дальнейшего удобства демонтажа.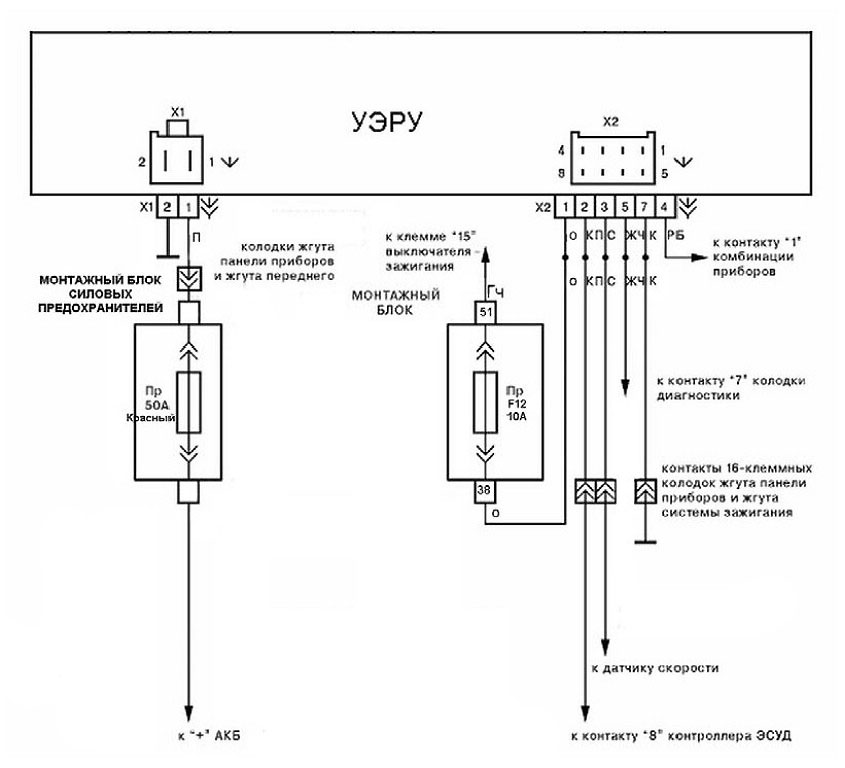 (ну мало ли что…)
(ну мало ли что…)Контрольную лампу я вывел на лампу «не пристёгнутый ремень», припаял провод, и для работы надо поставить светодиод. Эта лампа пригодится для самостоятельной диагностики неисправности ЭУР. (диагностика будет описана ниже)
После выводим провода на замок зажигания, подрулевых переключателей слева под панелью.
Соединяем их по схеме.
Ай чуть не забыл, теперь ведь можно на переключатель дворников, прилепить и задний. Тащим туда же провода с клавиши.
Подгоняем кожух рулевой колонки, запиливаем кнопку аварийки.(я её поставил справа от руля) Всё собираем, втыкаем предохранитель, который поставили под капот.
Запускаем двигатель и наслаждаемся лёгкому рулю. (контрольная лампа должна погаснуть после запуска)
PS Установил я датчик скорости, но трос на него не накручивается, мешает подушка.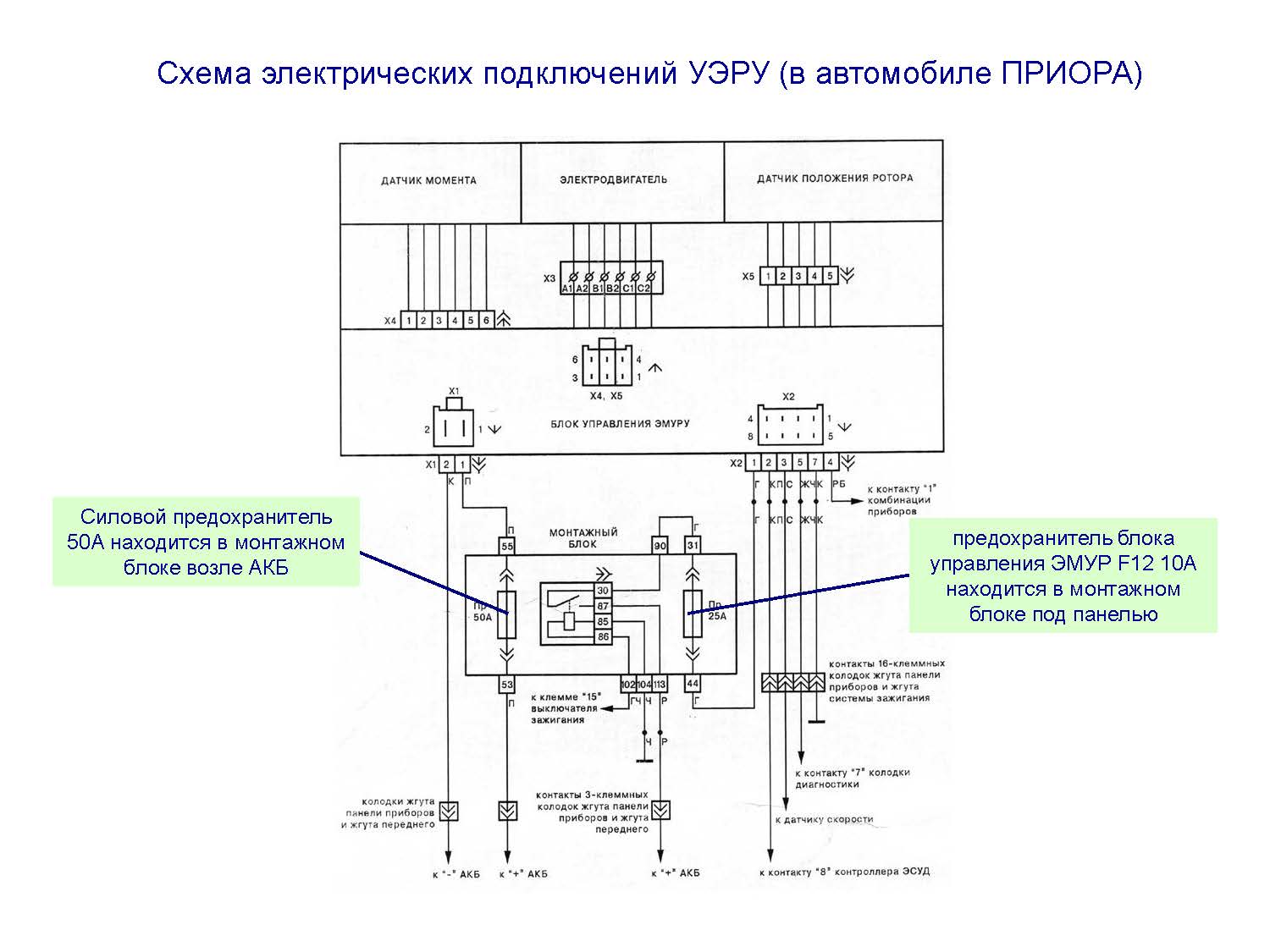 Поэтому пришлось купить приборку от чепырки, но это уже совсем другая история…
Поэтому пришлось купить приборку от чепырки, но это уже совсем другая история…
Ну и теперь не много слов про самостоятельную диагностику ЭУР. Для этого нужна канцелярская скрепка.
Пишу как делал лично я, а не как было написано на разных сайтах.
Вставляем скрепку в ЭУР в шестой пин, (он пустой, см. схему), так чтобы её можно было замкнуть на массу.
Запускаем двигатель, замыкаем на массу и отпускаем, контрольная лампа сначала моргнёт медленно (означает что ЭУР вошёл с систему диагностики). Далее лампа начнёт быстро моргать (считаем сколько раз моргнёт и см. расшифровку ниже
), после лампа опять медленно моргнёт (вышел из диагностики).— 1 Система исправна
— 2 Отсутствие сигнала оборотов двигателя
— 3 Неисправность датчика момента
— 4 Неисправность двигателя ЭМУР
— 5 Неисправность датчика положения рулевого вала
— 6 Неисправность датчика положения ротора двигателя ЭМУР
— 7 Неисправность бортсети автомобиля (ниже 10В либо больше 18В)
— 8 Неисправность блока управления ЭМУР
— 9 Неисправность датчика скорости автомобиля
(в источниках написано что лампа моргает на 10 раз больше, т. е. * 11 Система исправна и так далее, но у меня было так как я написал)
е. * 11 Система исправна и так далее, но у меня было так как я написал)
А начал я искать про диагностику, т.к. на ходу отключался ЭУР, а почему? Да потому что я не посмотрел надписи на датчике скорости, а подключил по логике, по цвету проводов на фишке от него, и получилось что перепутал полярность))) Поэтому и написал, что
Всем удачи и до встречи в новой записи)))
PS Если возникли вопросы или увидели ошибки, то говорите))
В автомобилях ВАЗ Приора электроусилитель руля выполняет функцию обеспечения более облегченного управления транспортным средством. С помощью ЭУР водитель может без проблем поворачивать рулевое колесо одной рукой. Какие неисправности характерны для усилителя и как произвести замену ЭУР в гаражных условиях — об этом мы расскажем ниже.
Возможные неисправности: признаки и причины
Способы определения механических повреждений ЭУР и рулевых механизмов
Самостоятельное снятие и установка усилителя
Фотогалерея «Снятие ЭУР»
Видео «Пособие по самостоятельному ремонту усилителя»
Комментарии и Отзывы
Возможные неисправности: признаки и причины
В каких случаях производится разборка и ремонт рулевой рейки, ЭУР и гидроусилителя, какова схема подключения усилителя, какие функции выполняет датчик момента?
Для начала давайте рассмотрим основные неисправности системы, почему она не работает и способы их устранения:
- Некорректная работа или поломка блока управления — одна из самых сложных причин неработоспособности ЭУР и ГУР в Ладе Приоре.
 При такой неполадки на экране бортового компьютера высветится соответствующее сообщение об ошибке. Если вы ее расшифруете, то узнаете о том, что компьютер зафиксировал неработоспособность управляющего модуля. Такие проблемы обычно связаны с неполадками в пайке.
При такой неполадки на экране бортового компьютера высветится соответствующее сообщение об ошибке. Если вы ее расшифруете, то узнаете о том, что компьютер зафиксировал неработоспособность управляющего модуля. Такие проблемы обычно связаны с неполадками в пайке.
Скорее всего, причина заключается в отсутствии соединения либо плохом контакте схемы усилителя с контроллером. Такая неисправность решается методом ремонта, то есть перепайки, либо замены, если речь идет об автомобиле на гарантии. Более точно узнать о неполадках можно путем проведения компьютерной диагностики. - Выход из строя контроллера скорости. При поломке датчика скорости электроусилитель самопроизвольно отключается или вовсе не работает, при этом на дисплее компьютера высветится ошибка о неработоспособности. В данном случае можно попытаться перенастроить девайс или поменять поврежденные участки электроцепи в месте соединения датчика с блоком управления. Иногда причина кроется в плохом контакте датчика с бортовой сетью, тогда разъем нужно будет просто поправить.
 Если неисправность связана с неработающим датчиком, то устройство подлежит снятию и замене.
Если неисправность связана с неработающим датчиком, то устройство подлежит снятию и замене. - Рулевое колесо проворачивается с большим трудом. В первую очередь причину следует искать в отсутствии смазки. Если масла в системе мало, то руль будет тяжело поворачиваться, при этом во время вращений может слышаться нехарактерный шум или скрип. Проверьте уровень смазки и добавьте масло при необходимости.
- Длительная эксплуатация электроусилителя с отсутствием смазки может привести к поломке насоса. Неисправность сопровождается нехарактерными для работы ЭУР звуками, а также трудным вращением руля. Если насос подлежит ремонту, то можно считать, что вам повезло, если же не подлежит, то устройство придется менять.
- Неисправностям в работе электроусилителя может способствовать и сниженное напряжение в электросети машины. Если напряжение в бортовой сети действительно скачет, то изначально следует произвести проверку генератора и электроцепи его подключения. Для диагностики рекомендуется использовать мультиметр или вольтметр (автор видео — Александр Логинов).

Способы определения механических повреждений ЭУР и рулевых механизмов
Проверка механических повреждений производится только тогда, когда ЭУР в целом функционирует некорректно, но на дисплее бортового компьютера нет ошибок. Для диагностики механических поломок следует повернуть руль в разные стороны до упора при запущенном моторе.
Ремонт ЭУР на Лада Приора. Снятие, установка и основные поломки
Давно прошли те времена, когда управление автомобилем напоминало комплекс силовых упражнений для рук. Сегодня для того, чтобы повернуть руль, достаточно легкого касания ладонью, и все за счет использования электроусилителя руля. Конечно же, это устройство усложняет конструкцию, а чем сложнее устроена любая, не только техническая система, тем в большей степени она подвержена сбоям и поломкам.
Каждая Лада Приора оснащается электроусилителем руля (ЭУР), и сегодня мы расскажем, как диагностировать возникшие с ним проблемы и провести его демонтаж без обращения в автосервис.
Оглавление:
1 Проблемы с ЭУР Лада Приоры: признаки и причины
1.1 ЭУР отказал полностью
1.2 Нестабильная работа руля
1.3 Стук в руле
2 Снятие и установка ЭУР
2.1 Как снять руль?
2.2 Крышка монтажного блока и подрулевые переключатели
3 Механические поломки ЭУР
4 Как сохранить подольше ЭУР в рабочем состоянии?
Проблемы с ЭУР Лада Приоры: признаки и причины
Предыдущие ВАЗовские модели снабжались гидроусилителем руля, который менее надежен, чем ЭУР. Тем не менее, электрический аналог этого устройства также подвержен возникновению неисправностей, которые нужно своевременно выявлять, иначе безопасность вождения будет под угрозой, не говоря уже о комфорте.
Во всех случаях, начиная движение, обращаем внимание на значок в виде руля с восклицательным знаком, который всякий раз вспыхивает на приборной доске, как только мы поворачиваем ключ зажигания. Если значок не гаснет, то это верный признак неполадки с усилителем.
ЭУР отказал полностью на Приоре
Бортовой компьютер при отказе ЭУР должен (как нам хотелось бы) высветить код неисправности устройства, которая привела к его отключению (коды ошибок, связанных с ЭУР, начинаются с литеры «С»).
Случается, что отказ ЭУР и спидометра происходят одновременно. Вышедший из строя датчик скорости не дает сигнала системе, которая управляет усилителем руля исходя из показателей скорости. При этом проблема может крыться в повреждении проводки, соединяющей спидометр, датчик скорости и ЭУР. Если с ней все в порядке, то следует заменить датчик скорости.

В других случаях причиной отключения ЭУР становятся такие факторы, влекущие падение напряжения в электросистемах автомобиля:
- загрязненные или пригоревшие контакты, нагар или окалина, препятствующие полноценной электропроводности;
- повреждения изоляции проводки;
- сбой в настойках блока питания;
- слабый заряд аккумулятора.
Если усилителю не хватает напряжения, то компьютерная диагностика покажет ошибку С1013. Для полноценной работы рулевого усилителя Приоры ему требуется минимум 13,5 В напряжения. Поэтому первичную диагностику проблем с напряжением следует повести таким образом:
- замерять напряжение на аккумуляторе. Возможно, его нужно просто зарядить;
- если с АКБ все в порядке, то проверяется изоляция проводки и контактов на предмет короткого замыкания.
Диагностировать повреждение проводов и контактов может автоэлектрик.
Нестабильная работа руля
Случается, что во время езды по ровной дороге руль ни с того ни сего заваливается в сторону.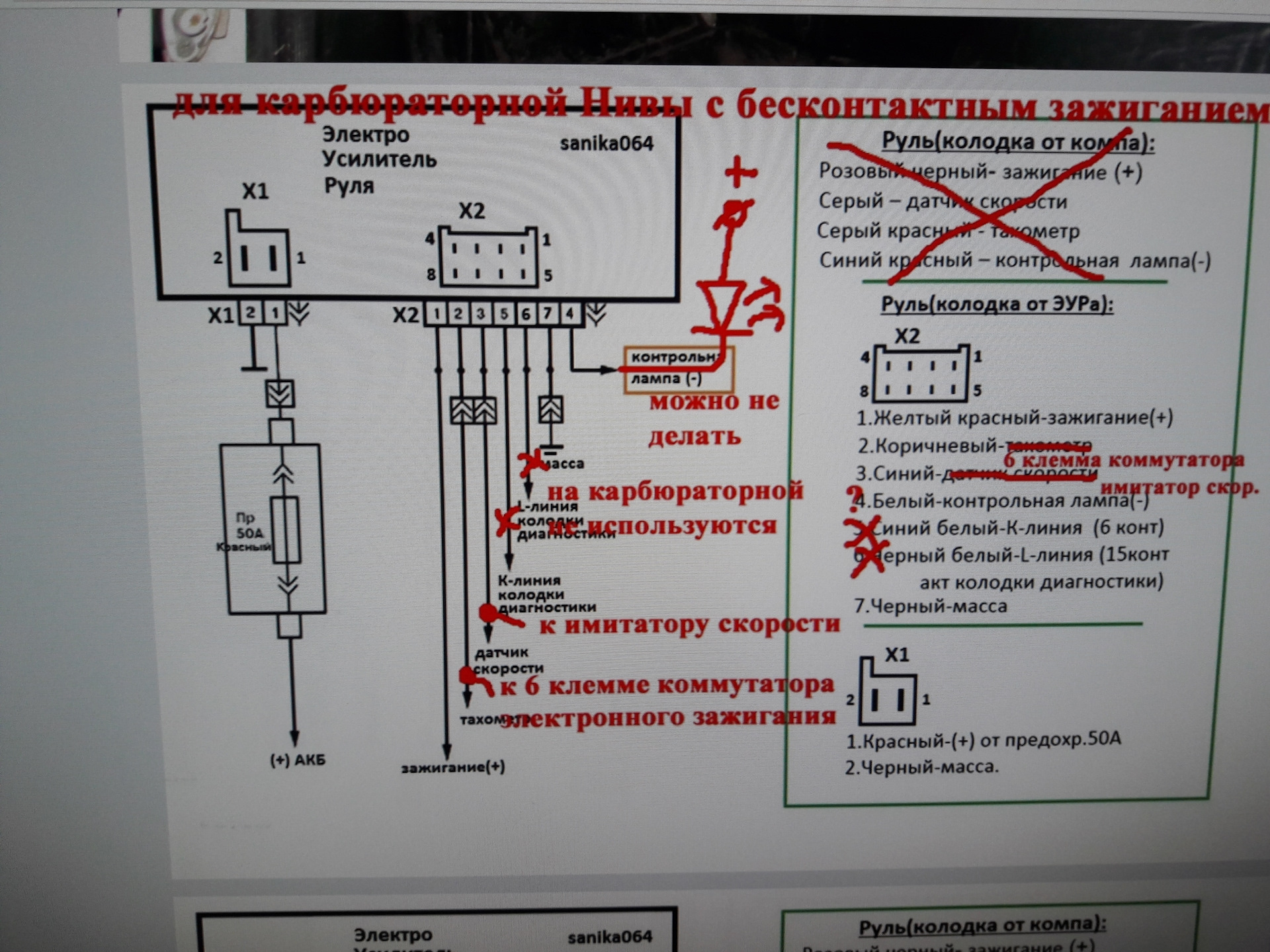 И такой вариант намного опаснее, чем полный отказ ЭУР. В последнем случае машина слушается, и можно по крайней мере добраться до СТО, в то время как при непредсказуемой работе усилителя в любой момент можно вылететь с трассы или выехать на встречку.
И такой вариант намного опаснее, чем полный отказ ЭУР. В последнем случае машина слушается, и можно по крайней мере добраться до СТО, в то время как при непредсказуемой работе усилителя в любой момент можно вылететь с трассы или выехать на встречку.
В таких случаях требуется полностью отключить усилитель, вытащив соответствующий предохранитель (F5) из монтажного блока салона. Как до него добраться, стоит почитать в отдельной статье. Сразу скажем, что проблема может крыться далеко и не только предохранителе, и чтобы понять, что на самом деле является причиной такой работы ЭУР, диагностику придется выполнить только на СТО, т.к. самостоятельно это сделать очень непросто.
Стук в руле на приоре
Чаще всего стук появляется из-за повреждений механической части рулевой системы (вала, рулевых реек и тяг, шаровых опор и т.д.). А вот на автомобилях Лада Калина и Гранта стук может быть связан с люфтом редукторного ЭУР, причем произведенного в Махачкале. Однако, если стук чувствуется и слышится на Приоре, то причину в ее рулевом усилителе нужно искать в последнюю очередь, поскольку автомобили ВАЗ 2170 редукторными, и тем более махачкалинскими ЭУР не комплектуются. А значит, обратить внимание надо на механическую часть ЭУР – вал, соединительные элементы, арматуру и другие детали, но уже после проверки остальных систем.
А значит, обратить внимание надо на механическую часть ЭУР – вал, соединительные элементы, арматуру и другие детали, но уже после проверки остальных систем.
В случае же механических повреждений ЭУР, бортовой компьютер может вообще не показывать ошибок, из-за чего приходится производить демонтаж и даже разборку усилителя.
Снятие и установка ЭУР
Чтобы оценить состояние механической части ЭУР или его контактов, требуется полностью извлечь его наружу. При этом собственно усилитель вытащить нетрудно, трудно получить к нему доступ. Для этого придется демонтировать руль, рулевой кожух и замок зажигания, крышку монтажного блока и целый ряд прочих узлов, интерфейс которых выведен на руль.
Обо всем этот по порядку.
Как снять руль на Лада Приора?
Для того, чтобы снять руль, нужно приготовить:
- торцовый ключ на 24 мм.;
- 5-мм. шестигранный ключ;
- плоскую и крестовую отвертки;
- молоток;
- зубило.
Кроме того, нужно обесточить автомобиль, отключив аккумулятор. Вообще этот пункт является обязательным при выполнении любых ремонтных работ. Дальнейший алгоритм представляет собой такую последовательность действий:
Вообще этот пункт является обязательным при выполнении любых ремонтных работ. Дальнейший алгоритм представляет собой такую последовательность действий:
- руль выставляется прямо;
- крестовой отверткой выкручиваем 7 шурупов, удерживающих кожух руля;
- находим замок зажигания и ищем его провода, где он соединяется с другими. Рассоединяем их;
- теперь предстоит снять замок зажигания, отвинтив четыре крепко закрученных болта. Для этого с помощью молотка и зубила наносят косые удары по головке каждого болта против часовой стрелки, как бы пытаясь их открутить. Может даже не получиться, поэтому часто приходится пользоваться дрелью, чтобы их высверлить. Вообще, болты эти повторно не используются, и щадить их не нужно, т.к. при каждой установке замка применяют новые болты;
- когда все 4 болта откручены, замок зажигания можно снять;
- с помощью 6-гранного ключа откручиваются 2 крепления подушки безопасности на руле, после чего саму подушку нужно немного провернуть;
- подушка отсоединяется от руля, если ее потянуть на себя и одновременно поддеть снизу плоской отверткой, однако тащить ее сразу нельзя: чтобы убрать ее в сторону, нужно поддеть и отвести фиксатор колодки проводов, которыми подушка сообщается с рулем;
- когда колодка проводов снята, подушку убираем в сторону;
- далее плоской отверткой отсоединяем колодку проводов, идущих на сигнал;
- теперь открылось крепление руля к валу.
 С помощью цветного маркера или другого обозначения, отмечаем положение рулевого колеса на валу, которое соответствует его прямой ориентации;
С помощью цветного маркера или другого обозначения, отмечаем положение рулевого колеса на валу, которое соответствует его прямой ориентации; - при помощи торцового ключа на 24 отдаем гайку, которой руль крепится на валу. При этом придется приложить силу, т.к. эта гайка затянута надежно;
- далее снимаем с вала руль, резко расшатывая или подбивая его из стороны в сторону, и после потягивая на себя.
Как видите, ничего сложного нет, кроме того, что требуются определенные физические усилия и грубые методы. Теперь дело за крышкой монтажного блока и подрулевыми переключателями.
Крышка монтажного блока и подрулевые переключатели
Вообще, релейно-предохранительных блоков на приоре три плюс один чисто предохранительный. Нас интересует крышка монтажного блока в салоне, который находится напротив левой ноги водителя.
Три замка крышки этого блока поворачиваются, в результате чего высвобождается нижняя часть крышки. Под ней мы находим рулевые переключатели, с помощью которых происходит указание поворотов, управление стеклоочистителями и связь с бортовым компьютером. Отключаются они следующим образом:
Отключаются они следующим образом:
- Пластиковые фиксаторы каждого (левого и правого) переключателя сжимаются;
- При сжатых фиксаторах каждый переключатель вынимается из гнезда;
- Выворачиваем 4 винта крепления усилителя приборной панели и снимаем этот усилитель.
- Отсоединяем колодку жгута проводов от катушки иммобилайзера.
Теперь остается заняться непосредственно усилителем руля.
Процесс изъятия электроусилителя
Для того, чтобы заняться непосредственно ЭУР, понадобятся головки на 8 и 13 мм., трещотка или вороток. Далее действуем следующим образом:
- С помощью 8-мм головки и трещотки отворачиваем контактную колодку и снимаем ее с вала;
- Отсоединяем все провода, ведущие к блоку управления ЭУР, обращая внимание на принцип работы проводных фиксаторов. Если с ним не разобраться, то фиксаторы можно повредить настолько, что потребуется замена этого элемента проводки;
- Находим карданный шарнир электроусилителя руля и головку стяжного болта на нем.
 Выкручиваем болт головкой на 13;
Выкручиваем болт головкой на 13; - С помощью молотка и зубила чуть расширяем стяжной хомут карданного соединения;
- Откручиваем 2 верхние гайки, которыми усилитель руля крепится к кузову;
- 2 нижние гайки не нужно откручивать полностью, достаточно провернуть их на 4 оборота, придерживая при этом ЭУР;
- После всего ЭУР можно, наконец, вынуть из кузова, сдернув с вала рулевой рейки.
Теперь у нас есть возможность полностью разобрать усилитель, произвести визуальную диагностику на предмет поломок, почистить и поставить обратно.
Установка отремонтированного или нового рулевого электроусилителя происходит в обратной последовательности. Перед этим важно проверить правильность расположения специальных пометок на защитном чехле, валу и крышке картера.
Конечно же, одно лишь снятие ЭУР не всегда дает возможность найти причину его неисправности, поскольку она может крыться в электротехнической части этого узла. В последнем случае понадобится тонкая диагностика на СТО, которую в гаражных условиях рядовой автомобилист вряд ли способен произвести.
Механические поломки ЭУР
Для чего, собственно, мы сняли ЭУР? Диагностика электротехнической части устройства не выявила проблем, а значит, нужно искать причину нарушений его работы в механике, которая может быть такой:
- Износ или засорение элементов привода или червячного механизма;
- Износ опорного подшипника червячного вала электропривода ЭУР;
- Другие признаки, указывающие на разрушение или износ деталей ЭУР.
В таких случаях требуется заменить изношенную деталь, по возможности попытавшись разобраться с факторами, повлиявшими на такой результат работы усилителя, или, в критическом случае – купить новый электроусилитель.
Как сохранить подольше ЭУР в рабочем состоянии?
Несмотря на то, что самостоятельно снять ЭУР можно и в гаражных условиях, без специальных инструментов и особых навыков, процедура эта небыстрая, т.к. к данному устройству быстро подобраться не получится. Поэтому автомобилистов интересует простой вопрос – как избежать необходимости ремонтировать или менять ЭУР. Ответ очевиден – нужно продлить срок его службы.
Ответ очевиден – нужно продлить срок его службы.
Для этого придется придерживаться таких правил:
- Никогда не трогаться с места, если руль находится в одном и крайних положениях. Надолго оставлять руль в таком положении тоже не рекомендуется даже без движения;
- Не удерживать руль дольше 4-5 секунд в крайних положениях, даже если того требует парковка или разворот. Лучше совершать нужные маневры;
- Обращать внимание на звуки, доносящиеся из электроусилителя;
- Прочищать контакты и подвижные механизмы от загрязнений;
- Обращать внимание на состояние проводки и клемм, если они попадаются на глаза в случае проверки других технических систем;
- Почаще проводить диагностику этого узла.
датчик момента, блок управления, схема, не работает
У современных автомобилей, в том числе и Лада Приора, руль можно крутить хоть двумя пальцами, даже в то время пока машина припаркована, раньше, если кто не знает, такого не было.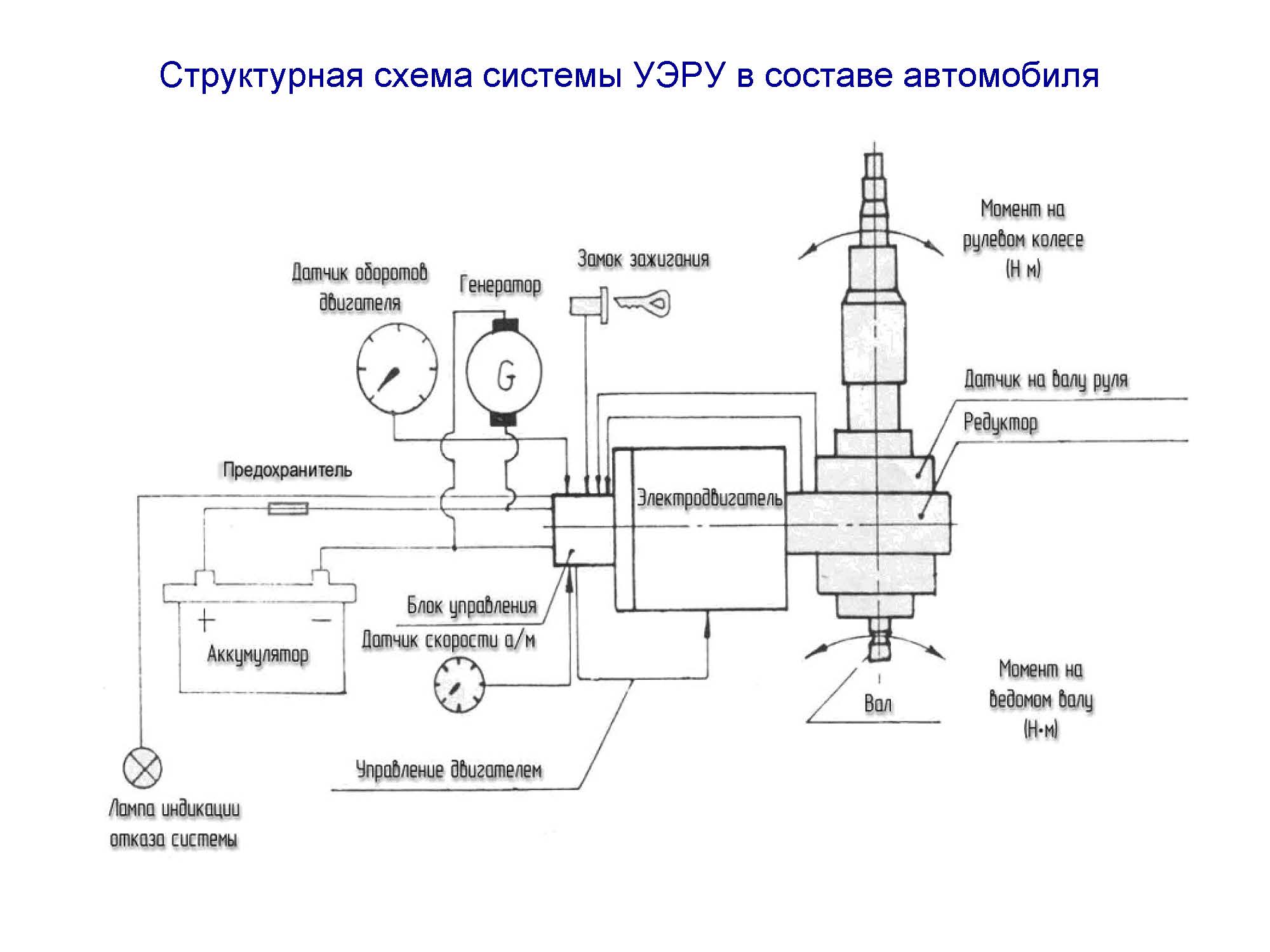 Благодаря современным технологиям появился электроусилитель руля. Однако, как многие знают, Приора, как и любое отечественное авто часто преподносит неожиданности, поэтому случаются ЭУР неисправности разного характера. В данном материале предлагаем разобраться, почему не работает такое крайне важное устройство, как электроусилитель руля и какие бывают способы устранения неприятностей связанных с этим.
Благодаря современным технологиям появился электроусилитель руля. Однако, как многие знают, Приора, как и любое отечественное авто часто преподносит неожиданности, поэтому случаются ЭУР неисправности разного характера. В данном материале предлагаем разобраться, почему не работает такое крайне важное устройство, как электроусилитель руля и какие бывают способы устранения неприятностей связанных с этим.
Почему ЭУР на Приоре лучше, чем ГУР
Благодаря ЭУР происходит более мощное усилие, передающееся в систему так сказать рулевого управления автомобилем. Если говорить простыми словами, то с таким устройством руль сидящий за ним, может крутить хоть 2-мя пальцами. Раньше на всех автомобилях представленных автомобилестроительной компанией «АвтоВАЗ» устанавливали ГУР, однако, с Калины от них руководством компании было решено отказаться, и перешли на более современные ЭУР.
Многие автовладельцы скажут, что гидроусилитель руля по своей мощности передачи намного предпочтительнее, однако, он имеет предостаточно недостатков, среди которых:
- Необходима регулярная проверка маслопроводов.

- Регулярно следует проверять и уровень в бачке.
- Силовой агрегат теряет в мощности.
Рассматриваемый нами сегодня электроусилитель руля, который установлен на Лада Приора, не имеет таких очевидных минусов, тем более в ремонте он прост, чем все тот же ГУР. Однако устройство не лишено, пусть и незначительных, но недостатков, случается даже так что, то работает то нет.
Как устроен электроусилитель рулевого колеса
Рассматриваемое нами в данном материале автомобильное устройство на Лада Приора, состоит из следующих компонентов:
- ЭБУ с соответствующей периферией. В этом блоке присутствуют разъемы со специальными датчиками.
- Электрическая часть. Данная часть является мотором устройства. Преимущественно этот компонент самый уязвимый.
- Механическая составляющая. Сюда включены элементы рулевого управления, крепеж и соединительные части. Этот компонент состоит: регулировочный рычаг, необходимый дабы сменить угол наклона рулевого колеса, а также выходной и входной валы, тут можно еще добавить пружины, присутствующие гайки, не пропустим шпильки и болты.

Определяем на Priora неисправности ЭУР
Самый главный признак ЭУР неисправности – когда срабатывает на приборной автомобильной панели лампочка, получающая сигнал от ЭБУ. В стандартной комплектации отечественной Лада Приора на «приборке» имеет такой значок, выполненный в виде колеса и восклицательного знака. Когда вы включаете в своем авто зажигание, то этот значок буквально на несколько секунд загорается, после чего гаснет, когда компьютер понимает, что все в порядке. Если же что-то не работает, то сигнал загорается и не перестает гореть до полного устранения неисправности.
Если лампочка горит, то самое верное решение в такой ситуации это пройти компьютерную диагностику, поскольку электроусилитель руля напрямую связан с бортовым компьютером Лада Приора. В настоящее время можно без труда приобрести диагностический индивидуальный сканер, которыми легко пользоваться и вы сможете легко и просто узнать причину неисправности, дабы самостоятельно выбрать способы устранения этой самой неисправности.
Итак, опытные водители рекомендуют провести диагностику, сделать это можно самостоятельно, имея под рукой соответствующее устройство. Вы сможете связаться с головным компьютером и получить все коды ошибок, причем неважно будут они без расшифровки или с ней. Если ваш сканер выдал вам лишь коды, не стоит отчаиваться, все легко можно узнать. Достаточно знать, что ЭУР неисправности обязательно будут начинаться с символа «С», к примеру, такой код как С1013 означает, что в сети слишком низкое напряжение, а ЭУР этого недостаточно.
Собственно при данной неисправности, вероятно, произошло ослабление, а возможно пригорание 1-го из 2-ух имеющихся силовых проводов, которые идут на усилитель руля. Провода там черный и красный большого сечения и входят они всегда в отдельный разъем на нашем устройстве, проверить их не составит труда. Также как вариант может быть неисправен предохранитель, с этой проблемой вообще все понятно и легко ее устранить. Еще одна причина, когда ЭУР не работает, это когда генератор или аккумулятор неисправен.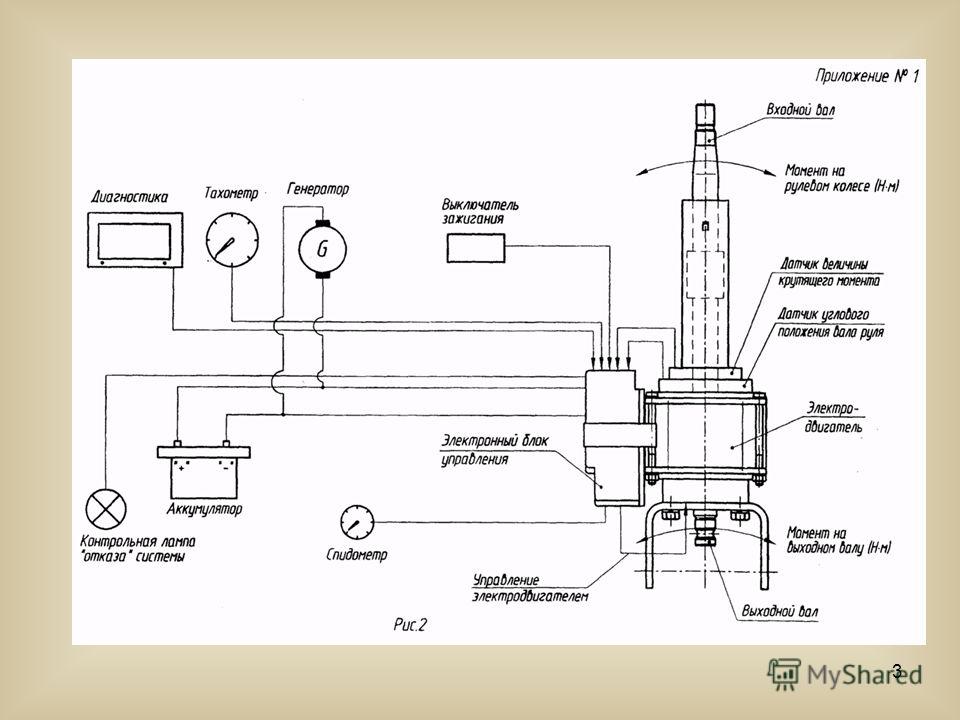 Если причина кроется в этом, то тут без хорошего специалиста электрика не обойтись никак.
Если причина кроется в этом, то тут без хорошего специалиста электрика не обойтись никак.
Противотуманные фары Лада Веста
Краш тест авторевю Веста
Лада Веста клиренс дорожный просвет
Используем «народный» метод проверки неисправности
В случае, когда на LADA Priora не горит лампочка, а ЭУР неисправности присутствуют, а возможно, требуется провести контроль, после проведенных ремонтных работ, то можно сделать так: покрутите автомобильный руль, без включенного зажигания, запомните при этом усилие. Далее запустите двигатель и повторите действие. Если нет никаких изменений между двумя действиями, то очевидно, что ЭУР не в порядке, нужна более детальная диагностика авто, после чего выбрать наиболее подходящие способы устранения неисправности, и это, скажем вам, совсем не проблематично.
Какие еще бывают неисправности и быстрые способы устранения
Вспоминаем опять составляющие ЭУР механика, электроника и соответственно электрика. С все той же механикой все более чем ясно и понятно, случается выход из строя компонентов: арматуры, вероятно соединений и/или валов. Если неисправность кроется в этом то, колеса вашей LADA Priora вообще не будут крутиться при движении рулевого колеса.
С все той же механикой все более чем ясно и понятно, случается выход из строя компонентов: арматуры, вероятно соединений и/или валов. Если неисправность кроется в этом то, колеса вашей LADA Priora вообще не будут крутиться при движении рулевого колеса.
В случае, когда неисправности кроются в другом, то дело обстоит сложнее. Когда удается понять, что из строя вышел усилитель или же сервомотор, то даже не стоит задумываться о ремонтных работах, в таком случае нужно приобрести новый усилитель и произвести замену. Если вы обнаружили, что между электроусилителем и автомобильным головным компьютером пропала связь, то причина кроется во 2-ом разъеме, то же самое, когда не поступает сигнал в блок от распределительного или коленчатого валов. Да, они крайне важны для работы устройства. В этой ситуации лучше отремонтировать или заменить провода.
Осталась элекронника устройства, и внутренние датчики положения валов. С этими самыми датчиками самостоятельно ничего сделать не получится, лучший вариант это ремонт в условиях автосервиса. Неисправность компьютера устраняется самостоятельно, вам необходимо купить его и установить.
Неисправность компьютера устраняется самостоятельно, вам необходимо купить его и установить.
Снимаем ЭУР на Приоре и устанавливаем новый
Когда ЭУР на вашем авто то работает то нет, и вы выявили причины, по которым понятно, что ремонтировать устройство бесполезно, нужно поменять усилитель на новый. Сделать это вы можете затратив немного времени и усилий, но все же своими руками.
Инструменты, необходимые для замены ЭУР на LADA Priora:
- Крестовая отвертка самая обыкновенная.
- Головка на «13», основной инструмент.
- Пригодится и ключ накидной на «8».
Пошаговая инструкция по замене ЭУР на Приоре:
- Первым делом обязательно требуется отключить аккумуляторную батарею, достаточно будет «скинуть» минусовую клемму.
- Снимаем с рулевой колонки пластиковые накладки, для этого отстегиваем пару фиксаторов и выкручиваем пару саморезов.
- Выкручиваем 4-е болта и отделяем свободно, но аккуратно нижнюю часть пластиковой закрывающей накладки.

- На данном этапе мешает замок и рычаги-переключатели понятное дело, дворников и поворотов. Не спеша, отключаем все провода и ключом на «8» требуется ослабить хомут, крепящий все выше перечисленные устройства. Они легко отходят с кожуха вала.
- Отсоединяем 2 разъема от электронного блока управления. Снимаем пластиковую накладку со специальной шпильки.
- Нижние гайки не нужно откручивать, достаточно повернуть их буквально на несколько оборотов.
- Далее, извлеките стяжной болт шарниров.
- Выкручиваем все оставшиеся крепежные гайки.
- Извлекаем ЭУР.
- Устанавливаем новый усилитель, и собираем все в обратной последовательности. Важно ничего не перепутать.
Подведем итоги
Все неисправности электроусилителя можно диагностировать самостоятельно на Приоре. Конечно же, не все неисправности вы сможете самостоятельно устранить, но замену устройства выполнить вполне реально, используя для этого пошаговую инструкцию.
схема и ремонт ЭУР и ГУР
В автомобилях ВАЗ Приора электроусилитель руля выполняет функцию обеспечения более облегченного управления транспортным средством. С помощью ЭУР водитель может без проблем поворачивать рулевое колесо одной рукой. Какие неисправности характерны для усилителя и как произвести замену ЭУР в гаражных условиях — об этом мы расскажем ниже.
Возможные неисправности: признаки и причины
В каких случаях производится разборка и ремонт рулевой рейки, ЭУР и гидроусилителя, какова схема подключения усилителя, какие функции выполняет датчик момента?
Для начала давайте рассмотрим основные неисправности системы, почему она не работает и способы их устранения:
- Некорректная работа или поломка блока управления — одна из самых сложных причин неработоспособности ЭУР и ГУР в Ладе Приоре. При такой неполадки на экране бортового компьютера высветится соответствующее сообщение об ошибке.
 Если вы ее расшифруете, то узнаете о том, что компьютер зафиксировал неработоспособность управляющего модуля. Такие проблемы обычно связаны с неполадками в пайке.
Если вы ее расшифруете, то узнаете о том, что компьютер зафиксировал неработоспособность управляющего модуля. Такие проблемы обычно связаны с неполадками в пайке.
Скорее всего, причина заключается в отсутствии соединения либо плохом контакте схемы усилителя с контроллером. Такая неисправность решается методом ремонта, то есть перепайки, либо замены, если речь идет об автомобиле на гарантии. Более точно узнать о неполадках можно путем проведения компьютерной диагностики. - Выход из строя контроллера скорости. При поломке датчика скорости электроусилитель самопроизвольно отключается или вовсе не работает, при этом на дисплее компьютера высветится ошибка о неработоспособности. В данном случае можно попытаться перенастроить девайс или поменять поврежденные участки электроцепи в месте соединения датчика с блоком управления. Иногда причина кроется в плохом контакте датчика с бортовой сетью, тогда разъем нужно будет просто поправить. Если неисправность связана с неработающим датчиком, то устройство подлежит снятию и замене.
- Рулевое колесо проворачивается с большим трудом. В первую очередь причину следует искать в отсутствии смазки. Если масла в системе мало, то руль будет тяжело поворачиваться, при этом во время вращений может слышаться нехарактерный шум или скрип. Проверьте уровень смазки и добавьте масло при необходимости.
- Длительная эксплуатация электроусилителя с отсутствием смазки может привести к поломке насоса. Неисправность сопровождается нехарактерными для работы ЭУР звуками, а также трудным вращением руля. Если насос подлежит ремонту, то можно считать, что вам повезло, если же не подлежит, то устройство придется менять.
- Неисправностям в работе электроусилителя может способствовать и сниженное напряжение в электросети машины. Если напряжение в бортовой сети действительно скачет, то изначально следует произвести проверку генератора и электроцепи его подключения. Для диагностики рекомендуется использовать мультиметр или вольтметр (автор видео — Александр Логинов).
Способы определения механических повреждений ЭУР и рулевых механизмов
Проверка механических повреждений производится только тогда, когда ЭУР в целом функционирует некорректно, но на дисплее бортового компьютера нет ошибок. Для диагностики механических поломок следует повернуть руль в разные стороны до упора при запущенном моторе. Разумеется, при этом машина должна находиться на одном месте. Если при вращении рулевого колеса вам приходится прикладывать значительно больше усилий, чем всегда, это говорит о неисправности в работе системы и поломке одного из компонентов. Такая же неполадка может свидетельствовать о выходе из строя других составляющих рулевой системы.
Для более точного определения неполадок следует осуществить тщательную проверку работоспособности. Вам нужно будет загнать свою Ладу Приору на эстакаду, яму либо подъемник, после чего внимательно произвести диагностику всех составляющих ходовой части. Нужно проверить рейки, рулевые тяги и прочие компоненты, поскольку есть вероятность, что сам усилитель рабочий, а признаки неисправности связаны с другими неполадками. Для обслуживания электроусилителя в домашних условиях вам придется полностью разобрать систему, очистить ее составляющие и собрать обратно.
Нужно проверить рейки, рулевые тяги и прочие компоненты, поскольку есть вероятность, что сам усилитель рабочий, а признаки неисправности связаны с другими неполадками. Для обслуживания электроусилителя в домашних условиях вам придется полностью разобрать систему, очистить ее составляющие и собрать обратно.
Схема распиновки и обозначений контактов ЭУР
Самостоятельное снятие и установка усилителя
Процедуру демонтажа и установки нового электроусилителя можно выполнить своими руками, но для этого вам надо будет приготовить все инструменты. Для выполнения работ вам потребуются две отвертки — одна с плоским наконечником, другая — с крестовым. Также понадобится и набор гаечных ключей с торцевыми головками.
Как выполнить задачу по снятию и установке своими руками:
- Перед выполнением работ нужно будет обесточить бортовую сеть вашей Приоры. Чтобы сделать это, выключите зажигание, а затем откройте капот и отключите аккумуляторную батарею от питания. После этого вам потребуется снять рулевое колесо, но перед этим демонтируйте пластмассовую накладку, расположенную под рулем и под приборной панелью.
 Кожух крепится на несколько болтов, их надо выкрутить и отложить в сторону. Снятие рулевого колеса осуществляется с учетом рекомендаций, описанных в сервисном мануале.
Кожух крепится на несколько болтов, их надо выкрутить и отложить в сторону. Снятие рулевого колеса осуществляется с учетом рекомендаций, описанных в сервисном мануале. - Когда накладка будет демонтирована вместе с рулем, вы увидите разъемы подрулевых переключателей, к которым подводятся провода. Эти штекеры нужно будет отключить. Также демонтажу подлежит и контактное кольцо.
- При помощи ключа с головкой на 8 вам потребуется выкрутить болт, который крепит разъем с проводами от выключателя зажигания. Отложите болт в сторону, чтобы не потерять.
- Когда эти действия будут выполнены, нажмите на пластиковые крепления и отключите разъемы с проводкой в верхней и нижней части рулевой колонки.
- Сделав это, в месте фиксации электрического усилителя непосредственно к колонке вы увидите пластиковую шторку. Вам нужно будет ее снять, для этого также потребуется отсоединить крепления.
- При помощи торцевого гаечного ключа вам надо будет незначительно ослабить гайки, с помощью которых крепится усилитель к нижней части колонки.
 Именно ослабить, а не выкручивать. Когда гайки будут ослаблены, нужно демонтировать электроусилитель из посадочного места, его снятие осуществляется с промежуточным валом. Но мы рекомендуем все же отсоединить усилитель от вала, это позволит быстро очистить ЭУР и упростить процедуру монтажа.
Именно ослабить, а не выкручивать. Когда гайки будут ослаблены, нужно демонтировать электроусилитель из посадочного места, его снятие осуществляется с промежуточным валом. Но мы рекомендуем все же отсоединить усилитель от вала, это позволит быстро очистить ЭУР и упростить процедуру монтажа.
Для отсоединения вала вам надо будет открутить винт, который крепит карданный шарнир на валу, с помощью гаечного ключа на 13. Сделав это, надо будет также выкрутить две гайки, расположенные на торцевых частях, после чего вся рулевая колонка подлежит снятию. - Затем вам надо будет демонтировать болт и кардан от шлицевого шкива механизма. При этом обратите внимание на метки — при дальнейшей установки они в любом случае должны совпадать. Метки расположены на защитном чехле, шкиве и крышке стартерного механизма. Теперь вы можете очистить электроусилитель и другие компоненты системы либо заменить вышедший из строя ЭУР на новый. Процедура сборки осуществляется в обратной последовательности.
Фотогалерея «Снятие ЭУР»
Видео «Пособие по самостоятельному ремонту усилителя»
Если вы столкнулись с необходимостью проведения ремонта электрического усилителя своими силами, то размещенный ниже ролик позволит вам без ошибок выполнить эту задачу (видео опубликовано каналом CompsMaster).
Понравилась статья? Поделиться с друзьями:
Снятие и установка ЭУР Приоры — «Клуб-Лада.рф»
| Электроусилитель руля Приоры (ЭУР) снимается вместе с рулевой колонкой для замены или ремонта. Установку рекомендуется выполнять вместе с помощником. |
Снятие электроусилителя руля ВАЗ 2170
Потребуется: Крестовая отвертка, накидной ключ «на 8», торцевой ключ «на 13».
Снять минусовую клемму с аккумулятора. Снять руль, декоративные накладки рулевой колонки и накладку панели приборов. Далее по порядку:
- Отвернуть 4 самореза с помощью крестовой отвертки, и снять нижнюю накладку панели.
- Отсоединить колодки с проводами от подрулевых переключателей и контактного кольца.
- Ослабить стяжной болт, используя накидной ключ «на 8».
 Разъединить колодки с проводами от замка зажигания.
Разъединить колодки с проводами от замка зажигания. - Сжать фиксаторы и отсоединить жгут с проводами от рулевой колонки.
- Сжать фиксаторы и отсоединить жгут с проводами в нижней части рулевой колонки.
- Освободить фиксаторы, и отсоединить 2 колодки с проводами от электроусилителя рулевого управления.
- Снять со шпильки крепления рулевой колонки декоративную шторку.
- Ослабить затяжку гаек нижнего крепления рулевой колонки, используя торцевой ключ «на 13». Отвернуть их на несколько оборотов.
Замечание:
| Снять рулевую колонку можно в сборе с промежуточным валом, но снимать и устанавливать колонку удобнее, если разъединить промежуточный вал. Соединение фланцев карданных шарниров промежуточного вала помечать не нужно, их установка возможна только в одном положении. |
Далее:
- Отвернуть гайку стяжного болта с помощью ключа «на 13» (он соединяет карданные шарниры промежуточного вала).

- Извлечь болт.
- Отвернуть левую гайку верхнего крепления рулевой колонки, используя торцевой ключ «на 13».
- Отвернуть правую гайку верхнего крепления рулевой колонки, используя торцевой ключ «на 13».
- Снять рулевую колонку Приоры в сборе.
- Отвернуть гайку стяжного болта фланца нижнего карданного шарнира и рулевого вала, используя ключ «на 13».
- Извлечь болт и снять карданный шарнир со шлицевого вала рулевого механизма.
Установка электроусилителя руля на Lada Priora
Установка рулевой колонки с ЭУР производится в обратной последовательности. Перед установкой убеждаемся, что паз на валу, метка на крышке картера рулевого механизма, и метки на защитном чехле расположены должным образом.
Расположение меток для установки рулевого вала:
1 — угловое углубление в крышке картера рулевого механизма; 2 — паз на валу рулевого механизма; 3 — крышка картера рулевого механизма; А — угловой выступ на защитном чехле; Б — вырез в кромке защитного чехла.
Проследите, чтобы метка А располагалась напротив углубления угловой формы (№1), которая расположена в крышке картера, а вал был повернут так, чтобы его паз (№2) располагался напротив метки Б.
Гайки стяжных болтов на фланцах промежуточного вала затягиваем моментом 23—28 Нм (2,3—2,8 кгсм).
Гайки крепления рулевой колонки затягиваем моментом 15—18,6 Нм (1,5-1,9 кгсм).
Устройство и схема электроусилителя руля Лада Приора
Элементы рулевого управления Lada Priora:
1 — правая рулевая тяга в сборе; 2 — правая опора рулевого механизма; 3 — скоба опоры рулевого механизма; 4 — промежуточный карданный вал; 5 — электроусилитель; 6 — рулевое колесо; 7 — труба рулевой колонки; 8 — левая опора рулевого механизма; 9 — рулевой механизм; 10 — левая рулевая тяга в сборе
Рулевая колонка с электроусилителем рулевого управления Приоры:
1 — входной вал; 2 — труба колонки; 3 — кронштейн колонки; 4 — электроусилитель; 5 — блок управления электроусилителем; 6 — рычаг регулировки наклона рулевой колонки; 7 — стяжная шпилька; 8 — задний кронштейн электроусилителя; 9 — пружина; 10 — выходной вал; 11 — болт-ось; 12 — передний кронштейн электроусилителя; 13 — силовой разъем блока управления; 14 — управляющий разъем блока управления; 15 — гайка.
Ключевые слова:
Понравилась статья? Поделитесь с друзьями!
диагностика неисправностей, монтаж и демонтаж
На автомобиле Лада Приора такое устройство, как электроусилитель руля, предлагается в стандартной комплектации, как функция комфорта. Именно об этом устройстве мы поговорим в этой статье. Предлагаем вам узнать, с какими неисправностями может столкнуться автовладелец этого авто и как произвести замену узла самостоятельно.
Содержание
[ Раскрыть]
[ Скрыть]
Роль электроусилителя в рулевом управлении ВАЗ 2170 Приора
Электроусилитель Приоры в снятом видеНи для кого не секрет, что электрический способ управления играет немаловажную роль в обеспечении безопасности во время езды. ЭУР или ГУР Приора выполняет опцию понижения уровня нагрузки на руль во время его поворота. Ключевым преимуществом использования такого устройства ЭУР на Приоре является комфорт при передвижении в результате улучшенной обратной связи между рулем и колесами.
Электроусилитель представляет собой более современный и усовершенствованный узел, который не только проще в плане использования, но и не требует внедрения дополнительных магистралей для обеспечения работы. Кроме того, данное устройство не забирает мощность у силового агрегата. Главное достоинство ЭУР — не требует частого ТО и контроля жидкости в отличии от ГУР.
Но следует отметить, что такая система имеет несколько недостатков, один из них — довольно слабая обратная связь между дорожным покрытием и системой управления. В результате этого при езде по снегу могут появиться трудности. В отличие от Приор, иномарки оснащаются дополнительными механизмами и узлами блокировки и стабилизации, поэтому способ управления ЭУР в отечественном варианте является менее надежным. Помимо этого, на практике неисправный электроусилитель с плохой тягой — это достаточно распространенное явление.
Схема подключения узлаРаспространенные неисправности и способы их устранения
С поломками устройств сталкиваются многие приороводы, в частности, речь идет о неисправности именно блока управления. Если блок управления системой работает некорректно или вышел из строя, на дисплее бортового компьютера появится соответствующая ошибка. Расшифровав ее, можно понять, что компьютер сообщает об отказе работы подключения ЭУР, что в большинстве случаев говорит о наличии проблем в пайке. В частности, речь идет о месте соединения платы усилителя с контроллером.
Если блок управления системой работает некорректно или вышел из строя, на дисплее бортового компьютера появится соответствующая ошибка. Расшифровав ее, можно понять, что компьютер сообщает об отказе работы подключения ЭУР, что в большинстве случаев говорит о наличии проблем в пайке. В частности, речь идет о месте соединения платы усилителя с контроллером.
Данная неисправность решается либо путем замены системы у дилера, если авто на гарантии, либо путем ремонта. Если у вас нет опыта в чип-тюнинге или работы с электронными девайсами, то решение такой неисправности необходимо доверить профессионалам. Разумеется, для того, чтобы точно поставить «диагноз», потребуется провести компьютерную диагностику.
Часто случается такое, что происходят сбои в работе датчика скорости, что опять же, способствует появлению ошибки бортового компьютера. Такой тип неисправности можно решить путем перепрограммирования устройства либо заменой проводов в тех местах, где регулятор соединяется с контроллером. Если сам регулятор является нерабочим, то его придется заменить (автор видео — Алексей a-skorp).
Если сам регулятор является нерабочим, то его придется заменить (автор видео — Алексей a-skorp).
Слишком низкий уровень напряжения в бортовой сети авто — это также распространенная проблема. В данном случае необходимо в первую очередь уделить внимание диагностике генераторного устройства либо проводки. Для проверки используйте тестер. В любом случае, если в работе системы обнаружены неполадки, необходимо произвести ремонт рулевой рейки своими руками. Эксплуатация авто с неработающим ЭУР не допускается до полного устранения проблемы.
Как определить механические повреждения ЭУР и рулевых механизмов?
Механическая диагностика осуществляется в том случае, если электроусилитель руля работает не нормально, но при этом бортовой компьютер не выдает ошибок. Чтобы произвести проверку механических повреждений, необходимо максимально провернуть рулевое колесо влево и вправо, мотор при этом должен быть заведен. Следует отметить, что при повороте рулевого колеса транспортное средство должно стоять на одном месте.
Следует отметить, что при повороте рулевого колеса транспортное средство должно стоять на одном месте.
В том случае, если вы заметили, что при повороте руля необходимо приложить больше усилий, чем обычно, то это может свидетельствовать о выходе из строя деталей или компонентов усилителя. Но также это может говорить о поломке других элементов рулевой системы (видео о диагностике усилителя руля с помощью скрепки представлено ниже, автор видео — Гоша Вахромеев).
Чтобы точно выявить, что же послужило проблемой, потребуется провести более тщательную диагностику. Для этого транспортное средство в первую очередь нужно загнать на подъемник и поднять, при этом тщательно проверив на работоспособность все элементы ходовки. Проверке подлежат рейки, рулевые тяги и т.д. Такую процедуру можно без проблем провести на любом СТО либо же своими силами, но только если у вас есть подъемник и определенные навыки по диагностике.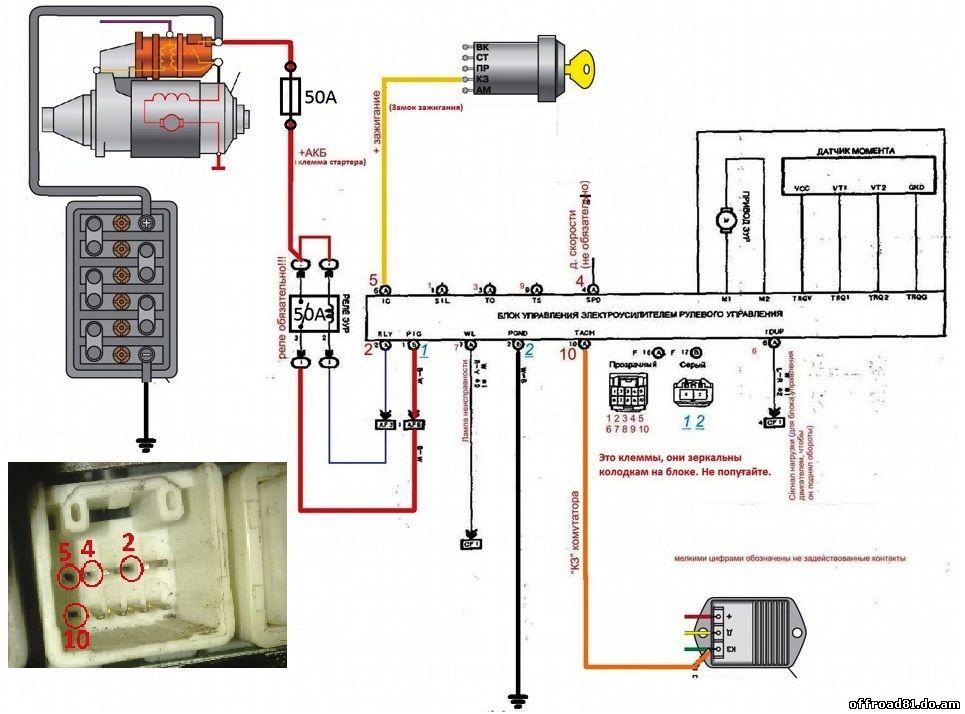 Есть вероятность того, что ремонт ЭУР не требуется, поскольку он сам по себе работоспособен, но требуется ремонт других элементов.
Есть вероятность того, что ремонт ЭУР не требуется, поскольку он сам по себе работоспособен, но требуется ремонт других элементов.
Чтоб не допустить появления неисправностей, связанных с электроусилителем, необходимо время от времени производить очистку системы, а также контактных узлов. Чтобы провести сервисное обслуживание самостоятельно, потребуется снять свой ЭУР, полностью разобрать его, произвести очистку и поставить обратно. Подробнее об этом мы расскажем ниже.
Снятие и установка ЭУР в домашних условиях
Для того, чтобы произвести снятие устройства с автомобиля, потребуется заранее подготовить все, что вам может пригодиться в процессе. В частности, речь идет об отвертках с плоским и крестовым наконечниками, а также наборе гаечных ключей с торцевыми головками.
Итак, как происходит процедура снятия и монтажа электроусилителя:
- В первую очередь, необходимо выключить питание на бортовую сеть, для этого отсоедините минусовую клемму от аккумулятора. Затем демонтируйте руль, заранее сняв пластиковую накладку под ним и на контрольном щитке.
 Демонтаж руля проводится в соответствии с сервисной книжкой к автомобилю.
Демонтаж руля проводится в соответствии с сервисной книжкой к автомобилю. - Используя крестообразную отвертку, необходимо открутить четыре болта фиксации декоративной накладки на рулевой панели.
- Сделав это, вы сможете увидеть колодки с проводкой — их необходимо отсоединить от подрулевого переключателя, также отсоединяется и контактное кольцо.
- Используя гаечный ключ на 8, нужно открутить винт фиксации колодок проводов, которые проходят от замка зажигания.
- Далее, необходимо нажать на пластмассовые элементы фиксации и отсоединить колодки с проводами вверху и внизу самой колонки.
- В том месте, где электроусилитель рулевого колеса крепится к самой колонке, потребуется демонтировать так называемую пластмассовую шторку, для этого следует отсоединить фиксаторы.
- Используя торцевой ключ, нужно несильно ослабить гайки, которые фиксируют узел к низу колонки.
После выполнения этих действий осуществляется снятие непосредственно усилителя — он демонтируется вместе с промежуточным валом. Но для того, чтобы вам было более удобно прочищать узел и в дальнейшем производить установку в обратном порядке, эти компоненты желательно заранее отсоединить друг от друга. Чтобы сделать это, используя ключ на 13 выкрутите винт, фиксирующий карданный шарниры на самом валу. Затем вам потребуется открутить две гайки фиксации по торцам, после этого саму рулевую колонку можно будет извлечь.
Но для того, чтобы вам было более удобно прочищать узел и в дальнейшем производить установку в обратном порядке, эти компоненты желательно заранее отсоединить друг от друга. Чтобы сделать это, используя ключ на 13 выкрутите винт, фиксирующий карданный шарниры на самом валу. Затем вам потребуется открутить две гайки фиксации по торцам, после этого саму рулевую колонку можно будет извлечь.
Затем нужно будет извлечь винт и произвести снятие кардана от шлицевого шкива узла. Чтобы после снятия процедура обратной установки была более удобной, желательно удостоверить в том, что все метки совпадают. Речь идет о метках на защитном чехле, шкиве, а также крышке стартера. Очистка механизма контактной группы и прочих элементов системы должна производиться осторожно, чтобы все компоненты остались целыми и неповрежденными. Если вы обратитесь за помощью к специалистам в этом вопросе, то процедура очистки обойдется вам в среднем около 30-50 долларов в зависимости от региона.
Загрузка …Видео «Установка усилителя своими руками»
Как правильно произвести монтаж электроусилителя в домашних условиях — смотрите на видео ниже (автор видео — sokur64).
Электроусилитель руля в карбюраторной Ниве
Электроусилитель руля в карбюраторной Ниве| Электроусилитель руля в
карбюраторной Ниве Автор Roadster4x4 |
Сообщения автора сделаны в конференции 06.10.03 (часть 1 и часть 2).
Отсканированные материалы присланы автором. Частично они повторяют уже выложенные в статье Электроусилитель руля, но имеют лучшее качество, частично — дополняют их.
[От Ведущего FAQ.
Хочу обратить внимание, что нынешнюю версию электроусилителя будет очень непросто установить на карбюраторную машину: он подключается к контроллеру и получает информацию о скорости и оборотах двигателя из него, а не непосредственно от датчиков: см. Электроусилитель руля — информация от разработчика.
ALER, 05.05.06
Смотрите также дополнение в конце этого файла, присланное mouse733/
ALER, 02.08.06]
Суббота — 6 часов установки
Воскресенье — тестовые заезды и регулировки
Приложенные документы:
Обозначение контактов
разъема
Схема монтажа жгута проводов
на ВАЗ-21214
Установка ЭУРУ на автомобиль
Схема крепления ЭУРУ и
кронштейна
Перечень кодов отказов
Особенности установки ЭУРУ
на автомобиль ВАЗ-21213
Установка
Установка была проведена в СПб в известном сервисе на Качалова, 9 известным мастером по спортивным Нивам Николаем Васильевым.
Машина — 21213 97 г. в. на резине 205/70R15 с дисками КРАМЗ 6.5″, вылет 40.
Дополнительно к комплекту ЭУР было привезено:
— РР с коротким валом;
— Крышка РК с двумя выходами (оказалось необязательно) с прокладкой и дополнительной шестерней;
— Датчик скорости (шестигранный) с ответной частью разъема.
Сейчас в продаже есть ДС с выходом на хвосте под тросик спидометра. В этом случае крышку РК с одним выходом менять не надо. ДС вкручивается в раздатку а на хвост ДС накручивается тросик спидометра.
Установка в основном аналогична описанной ранее установке на инжектор. Но есть некоторые отличия.
1. Заменить РР с длинным валом на РР с коротким. Говорят, бывают карбюраторные Нивы с коротким валом.
У меня была с длинным.
2. Заменить крышку РК или использовать ДС с выходом на тросик спидометра.
3. Изготовить кабель ДС. Я использовал три цветных провода, протянутых в кембрике. Длина кабеля
должна быть одинаковой с длиной тросика спидометра. Я делал 2 метра — лишнее отрезал по месту. Этим кабелем наращивается ответная часть разъема ДС. Распайка разъема указана в руководстве по установке.
В моем случае с прямоугольным разъемом последовательность контактов слева направо, если смотреть на торец разъема на ДС:
плюс — сигнал — земля.
4. Кабель ДС протягивается рядом с тросиком спидометра и заводится в салон через
то же отверстие в тоннеле. Под днищем кабель закрепили также за трос спидометра обыкновенными пластмассовыми электромонтажными хомутиками. Ушло 4 штуки.
5. Самая нетехнологичная операция — подсоединение провода к разъему коммутатора. Используется зеленый провод (крайний, ближайший к радиатору). Пришлось просто срезать часть изоляции и намотать сверху.
6. Плюс с замка зажигания я взял не из разъема на приборке, а с выхода 2-го предохранителя на длинной колодке. Это тот же самый провод. Я
переклинковал клемму, подсоединив дополнительный провод.
7. Все провода протягиваются под торпеду, и монтаж производится в отверстии под приборку.
8. Порядок соединения проводов:
— провода земли ЭУР и ДС на ближайшую гайку крепления воздуховодов;
— провода плюс ДС и оранжевый ЭУР соединяем с проводом с колодки предохранителей;
— сигнальный провод ДС соединяем с желто-голубым кабеля ЭУР;
— провод с коммутатора соединяем с розово-голубым кабеля ЭУР.
В разъемы приборки лезть мне не пришлось. При монтаже удобно иметь клинковщик (обжимщик клемм) и набор клемм с колпачками, ножевые папа-мама и с отверстием под болт.
Потом стандартная сборка.
Тестовые заезды и регулировка
Выехал из бокса — Вау! Развернуться в узком проезде в 5 приемов — не вопрос. Руль реально легкий. Проверено — 4-х летний ребенок на месте крутит руль от отбоя до отбоя.
Поехал домой — ощущение совершенно другой машины, надо полностью переучиваться ездить.
Для чистоты эксперимента в связи с заменой РР и просто для того, чтобы поставить руль ровно, заехал на сход-развал. Померили сход — в норме, выставили руль.
Мне показался великоватым люфт руля — вкрутил болтик на одну грань шайбы на РР, ослабил болты рулевого карданчика, покрутил руль, затянул.
Главный результат: ЭУР сохраняет постоянным усилие на руле на любых скоростях (0-100). До установки у моей машины был довольно острый руль с сильным эффектом возвращения руля. Причем на стоянке руль крутился довольно тяжело, на скорости — очень легко. Но эффект возвращения руля не позволял слишком сильно крутануть руль. После установки на всех скоростях руль стал нетяжелым, возвращающий эффект заметно ослаб. На стоянке руль вообще не возвращается. На скорости, например на выходе из поворота на разгоне, руль возвращается, но вяловато, или я просто не привык еще. Появилась возможность перекрутить руль в повороте — надо быть поосторожнее.
Важный момент — полностью исчез эффект увода в сторону на неровностях. Если раньше на неровной дороге приходилось довольно сильно подруливать, то сейчас просто ровно сидим и объезжаем ямки — как на мерседесе. Это может быть полезно гряземесам при вытаскивании из колеи, руль как известно там удержать очень тяжело. Надо будет попробовать еще на скорости на трассе с сильной колейностью.
В общем, ощущение что информативность руля пропала вообще. Просто крутим руль и колеса поворачиваются вне зависимости от внешних условий.
С выключенным усилителем ехать тяжело, но можно. Возвращающий эффект руля тоже заметно ослаб.
3.5 оборота это много, сюда бы РР с передачей поменьше.
Ездить очень удобно, посмотрим, как будет дальше когда привыкну. Но боюсь что в машину без усилителя я уже не сяду 🙂
Приложенные документы
06.10.03.
Дополнение от mouse733 — из документации на ЭМУР:
Особенности установки ЭМУР на а/м семейства 21213 (с карбюраторным двигателем).
1. Заменить крышку раздаточной коробки с одним гнездом на крышку с двумя гнездами (под трос спидометра и датчик скорости) 21214-3802824. Установить на эту крышку 6-тиимпульсный датчик скорости (например, 2112-3843010 или 2111-3843010).
2. Присоединить датчик скорости к жгуту ЭМУР. Распиновка находится внутри датчика скорости. +12 В (можно к оранжевому проводу комбинации приборов), сигнал (может быть обозначен буквой А) к серому проводу жгута ЭМУР и масса.
Коричнево-красный провод жгута ЭМУР присоединить к клемме 6 (зеленый провод) коммутатора.
02.08.06.
Рим — RacingCircuits.info
Информация о цепи
Адрес:
PH:
Тип схемы: Временный уличный курс
Веб-сайт: http://www.fiaformulae.com/en/championship/race-calendar/2020-2021/rome
История цепей
Circuito Cittadino dell’EUR в Риме — это временная трасса, созданная для чемпионата Формулы E, благодаря которой одноместные гонки проходят по улицам итальянской столицы.Это одна из самых длинных трасс, используемых электрическими гонщиками, и кульминация многолетних усилий, направленных на привлечение гонок в эту область.
Автодром находится в районе Esposizione Universale Roma (EUR) города. Район EUR был выбран итальянским диктатором Бенито Муссолини, чтобы прославить фашизм в 1940-х годах. Он надеялся организовать «всемирную ярмарку» на этом месте, но, хотя его амбиции вскоре были ограничены, огромный Колизей Квадрато, который также является фоном для трассы, был построен как символ фашистской архитектуры.Сегодня здесь расположены выставочные площади и штаб-квартира модного бренда класса люкс Fendi.
В своем первом воплощении трасса начиналась на Виа Кристофоро Коломбо, самой длинной районной дороге в Италии. Среди достопримечательностей — Обелиско ди Маркони, мраморно-бетонное сооружение, построенное в рамках проведения в городе Олимпийских игр 1960 года, и Римский конференц-центр (в котором находится E-Village во время гонок).
Первое мероприятие Rome E-Prix состоялось 14 апреля 2018 года перед 45 000 зрителей.Гонку с 33 кругами выиграл пилот Virgin Сэм Бёрд, занявший второе место на стартовой решетке. Лукас ди Грасси финишировал вторым для Audi, а пилот Techeetah Андре Лоттерер занял третье место.
В 2019 году второе издание гонки стало чем-то вроде триллера. Андре Лоттерер занял свою первую поул-позицию в сезоне, но на старте Хосе Мария Лопес вызвал огромное скопление в спине, в результате чего гонка была остановлена. Гонка была возобновлена под автомобилем безопасности, и Лоттерер лидировал, пока его не обогнал «Ягуар» Митча Эванса, который одержал свою первую победу для себя и команды.
В 2020 году повторения не должно было быть, поскольку гонка пала жертвой кризиса Covid-19, будучи одной из многих гонок, отмененных из-за пандемии.
В 2021 году гонка возвращается, но с существенно измененной компоновкой, которая призвана уменьшить влияние мероприятия на трафик за счет сокращения пробега по Via Cristoforo Colombo. Трасса протяженностью 2,1 мили с 19 поворотами является второй по длине трассой, когда-либо использовавшейся Formule E, разработанной при участии комитета гонщиков Лукаса ди Грасси, Жерома Д’Амброзио, Оливера Роуленда.
«Как всегда, мы создали настоящий вызов — и, на мой взгляд, это один из лучших макетов, которые мы когда-либо делали», — сказал спортивный директор Формулы E Фредерик Эспинос. «Первым и самым важным было создание пути, который вызвал бы минимальные неудобства для города, поэтому мы пересмотрели область, которую мы используем.
«Нам удалось сохранить характер Рима в этом новом макете с его волнами, множеством перепадов высот, изменений поверхности и волнением на трассе, которое мы всегда видели при посещении города.
«Рим является уникальным местом для уличной трассы в нашем календаре, так как это одна из немногих, где есть быстрые, широкие повороты и разные радиусы радиуса из-за дорог, с которыми нам приходится работать. Это схема, благодаря которой водители смогут реально изменить ситуацию ».
Обновленная планировка открывает доступ к римскому конференц-центру La Nuvola и создает пространство для большего количества зрителей на будущих мероприятиях. Реставрация тротуара на Виа Мурри — районе, имеющем историческое значение для города — и обновление дорожного покрытия вокруг трассы также укрепят наследие Формулы Е в Риме.
Как добраться
Circuito Cittadino dell’EUR находится в Риме, Италия. Аэропорт Леонардо да Винчи (во Фьюмичино) находится примерно в 20 минутах езды к западу, а аэропорт Чампино находится примерно на таком же расстоянии к юго-востоку.
Автодром находится в районе Esposizione Universale Roma (EUR) на юго-западе города. Есть множество вариантов общественного транспорта, чтобы добраться до этого района из центра Рима, в том числе метро, где станция EUR Palasport находится ближе всего к трассе, а трибуны и E-Village находятся в нескольких минутах ходьбы.
Проложить маршрут
Проезд и проживание
Найдите лучшие отели и предложения авиабилетов недалеко от Рима:
Возможно вам понравится:
Поделиться страницей
| Контрактная группа | 125 000 евро | ||||
| Часы торговли | CME Globex: | Воскресенье — Пятница 17:00м. — 4:00 дня. (18:00 — 17:00 по восточноевропейскому времени) с 60-минутным перерывом каждый день, начиная с 16:00. (17:00 по восточноевропейскому времени) | CME ClearPort: | Воскресенье 17:00 — Пятница 17:45 КТ без отчетов с понедельника по четверг с 17:45. — 18:00 CT | |
| Колебание минимальной цены | CME ClearPort: | 0,00001 за 1 евро с шагом 1,25 доллара США | |||
| CME Globex: | Итоги: 0.00005 за одно приращение в евро = 6,25 доллара Спреды за месяц подряд: 0,00001 за одно приращение в евро = 1,25 доллара Все другие комбинации спредов: 0,00002 за шаг за один евро = 2,50 доллара | ||||
| Код продукта | CME Globex: 6ECME ClearPort: EC Очистка: EC | ||||
| Перечисленные контракты | Квартальные контракты (март, июнь, сентябрь, декабрь), перечисленные на 20 кварталов подряд, и серийные контракты, перечисленные на 3 месяца подряд, | ||||
| Метод расчета | Срок поставки | ||||
| Прекращение торговли | Торговля заканчивается в 9:16.м. CT на второй рабочий день до третьей среды контрактного месяца. | ||||
| Порядок расчетов | Физическая поставка Процедуры расчетов по фьючерсам на EUR / USD | ||||
| Пределы позиции | Пределы положения CME | ||||
| Свод правил обмена | CME 261 | ||||
| Минимум блока | Минимальные пороги блока | ||||
| Предел цены или схема | Ценовые пределы | ||||
| Коды поставщиков | Цитировать Листинг символов поставщика | ||||
Circuito Cittadino dell’EUR — Схема проезда по Римской улице
Circuito Cittadino dell’EUR успешно дебютировал в календаре Формулы E в 2018 году и, по слухам, заключил долгосрочное соглашение о проведении электрических гонок в итальянской столице. Гонкапроводится на временной уличной трассе Circuito Cittadino dell’EUR, расположенной в районе EUR в Риме. С нее открывается великолепный вид на основные достопримечательности делового района, такие как Обелиско ди Маркони и Колизей Квадрато.
Трасса длиной 2,84 км является второй по длине трассой в календаре 5 сезона и известна своими волнистыми рельефами. Трасса с 21 поворотом, проходящим по склонам, является одной из самых ухабистых, что ставит перед водителями и командами различные задачи по управлению уровнем энергии автомобиля.Сэм Берд из Virgin Racing стал первым в истории победителем гонки Rome E-Prix, а титул лучшего круга в настоящее время принадлежит Даниэлю Абту (1: 37.910).
Rome не плохо знаком с уличными гонками и принимал множество других серий, прежде чем Формула E дебютировала там. На соседних улицах района EUR в 1920-х и 1930-х годах даже проходили Гран-при Рима.
Римская электронная премия 2020 года была отменена из-за пандемии COVID-19. Рим возвращается в календарь Формулы E в 2021 году, но Rome E-Prix 2021 года, который состоится 10 апреля, будет проходить за закрытыми дверями.
Узнайте больше о том, как добраться до Рима и Circuito Cittadino dell’EUR для участия в Rome E-Prix.Рим обслуживают два основных аэропорта, самый крупный из которых называется Леонардо да Винчи – Фьюмичино (FCO). Фьюмичино является крупным центром итальянского авиаперевозчика Alitalia и обслуживает рейсы в Италию, Европу, Северную Америку и Азию. Второй аэропорт — Чампино (CIA) — крупный хаб для Ryanair, из которого выполняются рейсы во все уголки Европы.
Circuito Cittadino dell’EUR расположен в муниципалитете X1 к югу от центра города.Он находится недалеко от нескольких станций метро (EUR Palasport и EUR Fermi) на южном конце синей линии метро B (см. Карту ниже). Вы также можете взять такси или прокатиться на трассе.
Схема метро Рима — нажмите, чтобы увидеть большую версию
Ознакомьтесь с нашими рекомендуемыми турами в Риме.
Представлен новый макет трассы для поездки Формулы E в Рим
Rome E-Prix, назначенный на 10 апреля, продемонстрирует впечатляющие исторические окрестности Римского квартала с пилотируемым полем полностью электрических одноместных автомобилей Формулы E 24 гонщика мирового класса примут участие в гонках перед Палаццо Конгресси, Пьяццале Маркони и, начиная с этого года, культовым Палаццо делла Цивилта Итальяна, также известным как «Площадь Колизея».Весь парк Нинфео также станет частью периметра трассы.
В результате тесного сотрудничества с местными властями был разработан план — при условии одобрения FIA — который сохраняет характер раунда и укрепляет наследие Формулы Е в этом районе.
ЧАСЫ: Rome E-Prix 2018/19, наш последний визит в столицу Италии
«Как всегда, мы создали настоящий вызов — и, на мой взгляд, один из лучших макетов, которые мы когда-либо делали», — сказал спортивный директор Формулы E Фредерик Эспинос.«Первым и самым важным было создание пути, который вызвал бы минимальные неудобства для города, поэтому мы пересмотрели территорию, которую мы используем.
«Нам удалось сохранить характер Рима в этом новом макете с его волнообразными волнами, множеством перепадов высот, изменений поверхности и волнением на трассе, которое мы всегда видели при посещении города.
«Рим является уникальным местом для уличной трассы в нашем календаре, так как он является одним из немногих с быстрыми, широкими поворотами и разным радиусом из-за дорог, с которыми нам приходится работать.Это схема, в результате которой водители смогут изменить ситуацию к лучшему.
«Мы показали планы гонщикам из нашего комитета (Лукас ди Грасси, Жером Д’Амброзио, Оливер Роуленд), и они были очень взволнованы, потому что это быстрый макет — очень быстрый — и это определенно сложно».
ESPORTS: Формула E: Accelerate продолжается, настраивайтесь каждый четверг в 19:00 по всемирному координированному времени
«Благодаря почти полностью обновленной схеме трассы в Риме, FIA и Formula E снова проделали потрясающую работу, сделав Формулу E еще более захватывающей. — сказал чемпион Формулы E 2017 года, представитель Audi Sport ABT Schaeffler Лукас ди Грасси.«Новая схема просто фантастическая: на ней более длинные и быстрые прямые, а также больше возможностей для обгона. Я уверен, что фанатам это понравится; Я не могу дождаться, когда поеду туда ».
Любое изменение схемы трассы дает спортивной команде Формулы Е возможность усложнить жизнь ее 12 высококлассным командам, инженеры которых гордятся созданием банков данных для максимального увеличения производительности своих автомобилей. «Скорость и эффективность на каждом E-Prix.
Эспинос говорит, что сочетание водителей и инженеров, которым необходимо настраиваться, а также характеристики трассы и возможности для обгона произведут настоящее зрелище для фанатов.
Цепь прямой связи регулирует выбор действий при ухаживании перед спариванием у самок дрозофил
Основные моменты
- •
Подмножество DA нейронов PPM3 снижает скорость копуляции девственной самки Drosophila 85
85 Цепь -FFL состоит из ГАМКергических и холинергических нейронов- •
ГАМК Рецепторы в нейронах ACh необходимы для подавления отторжения девственной самки
- •
Ретроградный сигнал NO, активируемый через глютти / NMD нейроны ГАМК
Резюме
На ранней стадии ухаживания самки плодовых мух проявляют острую реакцию отторжения, чтобы избежать неблагоприятного спаривания.Эта реакция отторжения перед спариванием эволюционно аналогична у разных видов, но молекулярная и нейронная основа такого поведения неясна. Здесь мы показываем, что предполагаемая некогерентная цепь прямой связи, включающая нейроны эллипсоидного тела, холинергический R4d и его репрессорные нейроны GABAergic R2 / R4m, регулирует реакцию отторжения перед спариванием у девственной самки Drosophila melanogaster . И R4d, и R2 / R4m положительно регулируются через специфические дофаминовые рецепторы подмножеством нейронов в дофаминергическом кластере PPM3.Генетическая депривация ГАМКергического сигнала через ГАМК Интерференция РНК рецептора в этой цепи вызывает массивный ответ отторжения, тогда как активация ГАМКергического R2 / R4m или подавление холинергического R4d увеличивает восприимчивость. Более того, глутаматергическая передача сигналов через рецепторы N -метил-d-аспартата индуцирует NO-опосредованную ретроградную регуляцию потенциально от R4d к R2 / R4m, вероятно, обеспечивая гибкий контроль поведенческого переключения от неприятия к принятию. Наше исследование проливает свет на молекулярные и нейронные механизмы, регулирующие поведенческий процесс отбора самок перед спариванием.
Ключевые слова
ухаживания
восприимчивость
сексуальная мотивация
дофамин
Drosophila
прямая связь
центральный комплекс
эллипсоидное тело
Рекомендуемые статьи
000 статьи
Ссылки на статьи
Подавление нисходящего префронтального контура предотвращает кетамин-индуцированную стрессоустойчивость у женщин
Резюме
Стресс является мощным этиологическим фактором в начале большого депрессивного расстройства и посттравматического стрессового расстройства (ПТСР).Поэтому были предприняты значительные усилия для выявления факторов, обеспечивающих устойчивость к последствиям более позднего стрессора, в надежде предотвратить неблагоприятные клинические исходы. Антагонист рецептора NMDA кетамин недавно появился в качестве профилактического средства, способного предотвращать нейрохимические и поведенческие последствия будущего стрессора. Несмотря на многообещающие результаты доклинических исследований, проведенных на самцах крыс, эффекты проактивного кетамина у самок крыс остаются неизвестными. Это вызывает тревогу, учитывая, что расстройства, связанные со стрессом, поражают женщин почти в два раза чаще, чем мужчин.Здесь мы исследуем профилактические эффекты кетамина на вызванное стрессом тревожное поведение и процессы на уровне нервных цепей, которые опосредуют эти эффекты у самок крыс. Кетамин, введенный за неделю до неконтролируемого стрессора (неизбежный удар хвостом; IS), снизил типичную вызванную стрессом активацию серотонинергического (5-HT) дорсального ядра шва (DRN) и устранил DRN-зависимый дефицит социального исследования подростков (JSE) через 24 часа после стрессор. Проактивный кетамин изменил нейронные ансамбли прелимбической коры (PL), так что более поздний опыт с IS теперь активировал эти клетки, чего обычно не происходило.Кетамин резко активировал цепь PL-DRN (PL-DRN) и ингибирование этой цепи с помощью дизайнерских рецепторов, исключительно активируемых дизайнерскими препаратами (DREADD) во время IS через неделю, предотвращало профилактику стресса, предполагая, что стойкие изменения в цепи PL-DRN Активность отвечает, по крайней мере частично, за опосредование долгосрочных эффектов, связанных с кетамином.
Заявление о значимости
Расстройства, связанные со стрессом, поражают женщин в два раза чаще, чем мужчин, поэтому выявление факторов, способствующих сопротивляемости женщин, имеет критически важное значение.Недавние усилия по выявлению нервных механизмов, лежащих в основе профилактического воздействия кетамина на поведенческое и нервное воздействие более позднего стрессора, были сосредоточены исключительно на грызунах-самцах. Здесь мы показываем, что кетамин, вводимый самкам крыс за неделю до неконтролируемого стрессора, предотвращает вызванные стрессом поведенческие эффекты, подобные тревоге. Были изучены механизмы, с помощью которых кетамин оказывает профилактическое действие, и было обнаружено, что кетамин активирует тормозную цепь предлимбической коры (PL) -дорсальное ядро шва (DRN), и что активация этой цепи необходима для смягчающих стресс эффектов кетамина.Эти данные служат основой для профилактического применения кетамина у женщин.
Введение
Психиатрические расстройства, связанные со стрессом, такие как депрессия и посттравматическое стрессовое расстройство (ПТСР), затрагивают женщин почти в два раза чаще, чем мужчины (Kessler et al., 2005; Steiner et al., 2005), и относятся к числу ведущие причины инвалидности во всем мире (Kessler et al., 1995; Mathers et al., 2008). Только одна треть пациентов, которым назначены обычные фармакотерапевтические препараты, достигают полной ремиссии, что подчеркивает необходимость более эффективных терапевтических методов.(Gaynes et al., 2009). Недавно было показано, что однократная субанестезирующая доза (0,5 мг / кг, внутривенно) неселективного антагониста рецепторов NMDA кетамина дает быстрые и стойкие терапевтические эффекты у людей с устойчивой к лечению депрессией, тревогой и посттравматическим стрессовым расстройством (Berman et al., 2000; Zarate et al., 2006; Glue et al., 2017; Price et al., 2009; Feder et al., 2014). Соответственно, все больше исследований посвящается выявлению основных нейробиологических механизмов, с помощью которых кетамин оказывает свое действие.
Из-за своей клинической эффективности лабораторная работа была сосредоточена на двух парадигмах. В одном случае однократная субанестезирующая доза (10 мг / кг, внутрибрюшинно) кетамина вводится в различные моменты времени перед поведенческими тестами, которые, как считается, отражают депрессивное или связанное с тревогой поведение. Например, кетамин, доставленный от нескольких минут до часов, прежде чем поведенческое тестирование предотвращает типичные поведенческие изменения, измеренные во время теста принудительного плавания (Garcia et al., 2008), теста подвешивания хвоста (da Silva et al., 2010), теста подавления новизны (NSF; Fuchikami et al., 2015) и тест в открытом поле (Thelen et al., 2016). Во втором случае кетамин назначают после воздействия стрессора, чтобы определить, сможет ли он обратить вспять влияние стресса на поведение. Кетамин, вводимый вскоре после (0–24 ч) воздействия хронического непредсказуемого стрессора, обращает действие стрессора на NSF и предпочтение сахарозы (Li et al., 2011). Удивительно, но почти все доклинические исследования, направленные на определение механистического действия кетамина, были сосредоточены на самцах крыс.Действительно, небольшое количество исследований продемонстрировало различную чувствительность и чувствительность женщин и мужчин к прямым и восстанавливающим эффектам кетамина (Carrier, Kabbaj, 2013; Frnasceschelli et al., 2015; Zanos et al., 2016).
В последнее время наблюдается большой интерес к факторам, которые могут привести к устойчивости перед лицом невзгод (для обзора см. Baratta et al., 2013), и, что интересно, 3 недавних отчета показывают, что однократная доза кетамина может притупить воздействие стрессоров, возникающих спустя две недели (Amat et al., 2016; Брахман и др., 2016; McGowan et al., 2017). К сожалению, ни в одном из этих отчетов не использовались женщины. Таким образом, мы решили изучить проактивные эффекты кетамина у самок крыс, а также основные процессы на уровне нервных цепей, которые опосредуют такие эффекты. Мы стремились определить, достаточно ли кетамина, доставленного за неделю до неконтролируемого стрессора (неизбежный удар хвостом; IS), для предотвращения тревожного поведения, измеренного во время социального исследования подростков (JSE) через 24 часа, аналогично тому, как это наблюдается у самцов крыс ( Amat et al., 2016). IS-индуцированные изменения поведения частично опосредуются активацией серотонинергических (5-HT) нейронов в ядре дорсального шва (DRN; Maier and Watkins, 2005). В частности, IS активирует 5-HT нейроны в средней и каудальной DRN (Grahn et al., 1999), что приводит к высвобождению 5-HT в областях проекции, которые являются проксимальными медиаторами вызванных стрессом поведенческих изменений, таких как базолатеральная миндалина (BLA; Amat et al., 1998; Christianson et al., 2010; Dolzani et al., 2016). Действительно, блокада рецепторов 5-HT2C в BLA устраняет снижение JSE, вызванное предшествующим IS (Christianson et al., 2010). Поэтому мы исследовали влияние кетамина на IS-индуцированную активацию Fos в DRN 5-HT нейронах, чтобы определить, снижает ли кетамин IS-индуцированную активацию DRN (Amat et al., 2016). Пластические изменения в прелимбической области (PL) медиальной префронтальной коры (mPFC), мощном регуляторе активности DRN (см. Обзор в Maier and Watkins, 2010), имеют решающее значение для буферного эффекта кетамина (Li et al. ., 2010; Lepack et al., 2016; обзор см. В Duman, Krystal, 2016). Таким образом, мы исследовали, изменяет ли кетамин нейронные ансамбли PL, так что более поздний IS теперь активирует те же самые ансамбли.Наконец, мы исследовали, активирует ли кетамин напрямую путь PL-DRN, и если да, то является ли активация пути PL-DRN критической для защитных эффектов кетамина во время более позднего ИИ.
Материалы и методы
Схема эксперимента
Первая серия экспериментов проверяла, может ли однократная доза кетамина смягчить поведенческие и нейрохимические эффекты ИИ. Таким образом, низкие дозы кетамина (10 мг / кг, внутрибрюшинно), защищающие от стресса у самцов крыс (Li et al., 2010; Amat et al., 2016) вводили самкам крыс за одну неделю (7 дней) до лечения IS или HC. Отдельные группы крыс получали кетамин в высоких дозах (40 мг / кг, внутрибрюшинно), не влияющий на устойчивость к стрессу (Chowdhury et al., 2017), или физиологический раствор. Тревожное поведение оценивали во время теста JSE через 24 часа после воздействия стрессора. Таким образом, эксперимент представлял собой факторный план 2 (стресс) × 3 (лекарство). Для статистического анализа использовали двусторонний дисперсионный анализ. Предыдущая работа, выполненная с использованием аналогичных параметров (Dolzani et al., 2016) показывает, что n = 9–12 / группа достаточно для достижения статистической значимости между группами. Три крысы считались статистически выпадающими (> 2,5 SD от среднего) и были исключены из статистического анализа. Период времени в одну неделю был выбран, чтобы отделить долгосрочные буферные эффекты кетамина от стресса от потенциальных острых эффектов, которые не являются предметом внимания настоящей работы. Кроме того, предыдущая работа нашей лаборатории демонстрирует, что кетамин, введенный за одну неделю до IS у самцов крыс, защищает от типичных эффектов стрессора (Amat et al., 2016). Активацию DRN 5-HT оценивали в отдельной группе крыс с использованием иммуногистохимии с двойной меткой (IHC), чтобы определить, снижает ли кетамин общую стресс-индуцированную экспрессию Fos в DRN и происходит ли это снижение в нейронах 5-HT в DRN. Экспрессию Fos и 5-HT исследовали в ростральной, средней и каудальной DRN. Таким образом, эксперимент представлял собой факторный план 2 (стресс) × 2 (лекарство). Двусторонний дисперсионный анализ использовали для отдельного анализа общего количества 5-HT + клеток, общего Fos + и процента 5-HT + клеток, которые также были Fos +, между всеми возможными группами.Предыдущая работа нашей лаборатории (Dolzani et al., 2016) демонстрирует, что n = 10–12 / группа (два среза мозга на крысу) достаточно для обнаружения статистических различий в иммуногистохимической маркировке между группами. Четыре крысы считались статистически выпадающими (> 2,5 SD от среднего по группе для количества клеток или недостаточного окрашивания для обнаружения) и были исключены из анализа.
Главный источник ингибирующего контроля над нейронами DRN 5-HT происходит от нисходящих глутаматергических пирамидных нейронов PL, которые синапсируются преимущественно на интернейронах GABA в DRN (Jankowski and Sesack, 2004).Интересно, что поведенческий контроль притупляет воздействие стрессора, активируя этот тормозной путь (Amat et al., 2005; Baratta et al., 2009). Более того, опыт контроля также имеет профилактический эффект, поскольку он блокирует поведенческие эффекты более поздних неконтролируемых стрессоров, таких как IS (Amat et al., 2006) и социальное поражение (Amat et al., 2010). После предыдущего опыта контроля неконтролируемый стрессор теперь не активирует DRN 5-HT нейроны (Baratta et al., 2009). Это происходит потому, что опыт контроля изменяет путь PL-DRN, так что теперь он активируется даже неконтролируемыми стрессорами, такими как IS (Baratta et al., 2009). Таким образом, вторая серия экспериментов была направлена на то, чтобы определить, может ли кетамин задействовать те же механизмы, что и поведенческий контроль, и изменять нейроны PL, так что более поздний IS теперь активирует PL. Мы использовали недавно разработанную платформу Robust Activity Marker (RAM) для немедленного раннего гена (IEG), чтобы исследовать нейронную активность в двух временных точках: время первоначальной инъекции кетамина (время 1) и время более позднего IS (время 2). RAM предоставляет средства для маркировки нейронных ансамблей, активированных конкретным временным переживанием или событием (подробное объяснение см. В Sørensen et al., 2016). В сочетании с последующей техникой маркировки IEG, такой как IHC, RAM позволяет запрашивать нейронную активность в нескольких временных точках. RAM использует синтетический промотор, зависящий от активности (pRAM), который управляется нейрональной специфической активностью FOS и NPAS4. Активность pRAM управляет экспрессией домена трансактиватора тетрациклина (tTA), который связывается с элементом ответа на тетрациклин (TRE) и управляет экспрессией эффекторного гена mKate2. Временной контроль над ОЗУ достигается с помощью модифицированной системы Tet-Off.Связывание tTA с TRE ингибируется в присутствии DOX (DOX +). В отсутствие DOX (DOX-) активируется транскрипция эффекторного гена. Поэтому мы использовали RAM, чтобы определить, активирует ли кетамин нейроны PL во время инъекции (время 1) и активирует ли более поздний опыт IS (время 2) теперь те же или другие нейронные ансамбли, как и предыдущий кетамин. Важно отметить, что кетамин в низких и высоких дозах вводили отдельным группам крыс, чтобы определить, является ли IS-индуцированная активация нервных ансамблей, ранее активированных кетамином, специфической для дозы кетамина, которая защищает от поведенческих исходов стресса.Таким образом, эксперимент представлял собой факторный план 2 (стресс) × 3 (лекарство). Двусторонний дисперсионный анализ был использован для отдельного анализа общего RAM, общего Fos + и процента RAM +, которые также были Fos +, между всеми группами. Предыдущая работа Соренсена и др. (2016) показывает, что n = 6–8 на группу (два среза мозга на крысу) достаточно для выявления статистических различий между группами в экспериментах, проводимых с использованием аналогичных параметров. Пять крыс считались выбросами (> 2,5 SD от среднего количества клеток или неспособность определить экспрессию инъекционного контроля (eYFP).
В третьем эксперименте проверяли, активирует ли кетамин цепь PL-DRN. Красные флуоресцентные ретроградно транспортируемые микросферы (далее именуемые ретробусами; RB) вводили в DRN за две недели до того, как крысы получали однократную инъекцию кетамина. Два часа спустя крыс умерщвляли и оценивали экспрессию Fos в путях PL и PL-DRN. Тест независимых образцов t был использован для исследования различий в общем количестве RB-положительных (RB +) клеток, общем количестве клеток Fos + и процентном содержании клеток RB +, которые также были Fos +; n = 6–8 на группу (два среза мозга на крысу) достаточно для выявления статистических различий между группами.Две крысы считались выпадающими (> 2,5 SD от среднего количества клеток) и исключались из анализа. Кроме того, 4 крысы были исключены из анализа из-за неточных инъекций DRN RB.
Последний эксперимент проверял, необходим ли контур PL-DRN для защитных эффектов кетамина. Это требует избирательного ингибирования нейронов PL, которые проецируются в DRN. Решение этой проблемы потребовало использования двойной вирусной межсекционной генетической стратегии для нацеливания рецепторов-конструкторов, активируемых исключительно дизайнерскими препаратами (DREADD), на нейроны PL, проектирующие DRN.Ретроградно транспортируемый вектор AAV, кодирующий рекомбиназу Cre, доставляли в DRN, и дважды флоксированный вектор AAV, кодирующий ингибирующий рецептор DREADD (hM4Di) или mCherry (контрольный вирус), доставляли в PL. Этот подход позволяет избирательно ингибировать нейроны после системной инъекции клозапина- N -оксида (CNO; Armbruster et al., 2007; Ferguson et al., 2011; Rogan and Roth, 2011), что подтверждается сниженной экспрессией c -Fos (Ferguson et al., 2011; Soumier and Sibille, 2014).Чтобы подтвердить опосредованное hM4Di ингибирование пути PL-DRN, мы использовали тест независимых образцов t для оценки общей экспрессии PL-DRN hM4Di / mCherry, общего Fos + и процента клеток, экспрессирующих hM4Di / mCherry, которые также были Fos +. В отдельной когорте крысам, экспрессирующим hM4Di или mCherry в пути PL-DRN, вводили кетамин или физиологический раствор, а через неделю они получали инъекцию CNO за 30 минут до IS. Тревожное поведение оценивали через 24 часа во время JSE. Таким образом, эксперимент представлял собой факторный план 2 (вирус) × 2 (стресс) × 2 (лекарство).Для статистического анализа использовали трехфакторный дисперсионный анализ. Предыдущая работа с использованием аналогичных параметров (Dolzani et al., 2016) демонстрирует, что n = 10–12 / группа достаточно для выявления статистической значимости. Три крысы считались статистически выпадающими (> 2,5 SD от среднего) и были исключены из статистического анализа. Кроме того, две крысы были исключены из-за травм лап, связанных со стрессом, и шесть крыс были исключены из-за неточных или неудачных инъекций вируса, как было определено после завершения эксперимента.
Анализ данных для всех тестов однофакторного дисперсионного анализа, двустороннего дисперсионного анализа и тестов t проводили с использованием программного обеспечения Prism (GraphPad). Трехфакторный дисперсионный анализ выполнялся с использованием Statview (SPSS). Все эксперименты проводились с использованием межгруппового дизайна, а эффект лечения анализировался с помощью непарного теста t (лекарство), одностороннего (стресс), двухстороннего (стресс и лекарство) или трехстороннего (стресс и наркотики). препарат и вирус) ANOVA. Основные эффекты и взаимодействия считались статистически значимыми, если p <0.05. При необходимости, было выполнено апостериорных анализов и запланированных сравнений с использованием метода Тьюки апостериорных . Значения на графиках представлены как среднее ± SEM.
Крысы
Взрослых самок крыс Sprague Dawley (250–300 г; Envigo) содержали парами при 12/12 часовом цикле свет / темнота (свет включается в 7 утра и выключается в 7 вечера). Крыс содержали в условиях свободного доступа к пище и воде, и им позволяли акклиматизироваться к условиям колонии в течение 7 дней перед хирургическими или экспериментальными процедурами.Все стереотаксические операции выполнялись под анестезией изофлураном 2,5% (Piramal Critical Care). Перед операцией крысам вводили анальгетик (мелоксикам, 0,5 мг / кг, подкожно; Ветмедика) и антибиотик (Комби-Пен-48, 0,25 мл / кг, подкожно; Бимеда). Перед экспериментом крысам дали две недели на восстановление после операции. Все эксперименты проводились между 9 часами утра. и 17:00. Все процедуры с животными были одобрены Комитетом по уходу и использованию животных в Университете Колорадо в Боулдере и соответствовали рекомендациям Национального института здравоохранения по уходу и использованию лабораторных животных.
Стресс-процедура
IS была проведена, как описано ранее (Amat et al., 2005; Christianson et al., 2013). Вкратце, крыс помещали в удерживающую трубку PlexiGlas (8 × 18 см, диаметр × длина). Хвост крысы был прикреплен к опоре из оргстекла, выступающей из задней части бокса, с помощью медицинской ленты, а медные электроды были помещены вокруг хвоста. Электрошок прикладывался к хвосту крысы с возрастающей интенсивностью по мере развития сеанса электрошока (33 испытания при 1,0 мА, 33 испытания при 1 мА).3 мА и 34 попытки при 1,6 мА). Разряд производился со средним интервалом между испытаниями (ITI) 60 с. Крыс извлекали из боксов PlexiGlas и помещали в их домашнюю клетку сразу после последнего удара хвостом. Контрольных крыс, не подвергавшихся электрошоковой обработке, оставляли в колонии нетронутыми.
JSE
Тестирование JSE проводилось через 24 часа после IS или HC, как описано ранее (Amat et al., 2016). Любые крысы, показывающие признаки травмы (включая поврежденные задние лапы, передние лапы или ногти на ногах) после стрессового сеанса, исключались.Принимаются меры предосторожности, чтобы свести к минимуму частоту этих случаев, но, тем не менее, травмы произошли у небольшой группы пациентов. Каждому испытуемому по отдельности приписывали пустую пластиковую клетку с подстилкой из дерева и проволочной крышкой. Подопытные оставались в испытательной клетке от 45 минут до 1 часа перед тем, как ввести молодую (28 ± 2 дня) самку сородича. Наблюдатель, слепой к лечению, регистрировал исследовательское поведение (аллогруминг, облизывание, обнюхивание и приклеивание), инициированное подопытным в течение 3-минутного теста.Результаты теста JSE были представлены как общее время (а) социального исследования в течение 3-минутного теста.
Флуоресцентный IHC
Окрашивание на Fos проводили с использованием общего протокола иммунофлуоресценции. После серии промываний в 0,01 M PBS, содержащем 0,5% Triton X-100, срезы инкубировали в течение ночи в блокирующем растворе PBS, содержащем 0,5% Triton X-100 (PBST) и 2,5% бычий сывороточный альбумин (BSA), при 4 ° C. Затем срезы промывали PBST и инкубировали в течение 24 ч при комнатной температуре (RT) с кроличьими поликлональными первичными антителами (1: 1000; Santa Cruz Biotechnology) в блокирующем растворе.После серии промывок PBS срезы инкубировали в течение 2 ч при комнатной температуре в козьих вторичных антителах против кролика Alexa Fluor 405 (1: 250; Life Technologies). После серии промывок PBS ткань переносили на предметное стекло и покрывали покровным стеклом Vecta Shield (Vector Labs).
Анализ изображений для флуоресцентных экспериментов IHC
Срезы мозга наблюдались с помощью лазерного сканирующего конфокального микроскопа высокого разрешения со структурированным освещением Nikon N-SIM (Nikon). Изображения были получены с помощью NIS Elements (Nikon) и проанализированы с помощью FIJI (ImageJ).Все цифровые изображения были получены с использованием объектива 20x. Для визуализации и количественной оценки PL (взятых между AP: +2,5 мм до +3,0 мм относительно брегмы) границы были основаны на ранее описанных (Baratta et al., 2009) с учетом атласа головного мозга (Paxinos and Watson, 2011) . В эксперименте по ретроградному отслеживанию PL-DRN тельца клеток RB + в PL наблюдались с использованием лазерной линии с длиной волны 546 нм и были окрашены в красный цвет. Ядра Fos + наблюдались с помощью лазерной линии с длиной волны 405 нм и были окрашены в синий цвет.RB + и Fos + клетки в PL были количественно определены и записаны отдельно. Сообщалось о совместной локализации RB и Fos, когда наблюдали и проверяли, что красное / пурпурное тело клетки, представляющее смесь двух флуорофоров, перекрывает клетки RB + и Fos +. В эксперименте PL RAM нейроны, меченные eYFP (место вирусной инъекции), наблюдались с помощью лазерной линии с длиной волны 488 нм и были окрашены в псевдо-зеленый цвет. Экспрессия eYFP была подтверждена у всех субъектов, включенных в статистический анализ. После подтверждения точной вирусной инъекции ядра, меченные RAM, наблюдались с помощью лазерной линии с длиной волны 546 нм и были окрашены в красный псевдоцвет.Ядра Fos + наблюдались с помощью лазерной линии с длиной волны 405 нм и были окрашены в синий цвет. Клетки RAM + и Fos + в PL были количественно определены и записаны отдельно. Сообщалось о совместной локализации RAM и Fos, когда наблюдали и проверяли, что пурпурное ядро, представляющее смесь двух флуорофоров, перекрывает RAM и Fos. В эксперименте по подавлению опосредованного DREADD пути PL-DRN положительные клетки DREADD или mCherry наблюдались с использованием лазерной линии с длиной волны 546 нм, и они были окрашены в красный цвет. Клетки Fos + наблюдались с использованием лазерной линии с длиной волны 405 нм и были окрашены в зеленый псевдо-цвет.Клетки DREADD + и Fos + в PL были количественно определены и записаны отдельно. Сообщалось о совместной локализации DREADD и Fos, когда наблюдали и проверяли, что желтое ядро, представляющее смесь двух флуорофоров, перекрывает DREADD и Fos. Для каждого субъекта были взяты два отдельных подсчета для ткани PL, охватывающей AP: от +2,5 мм до +3,0 мм. Два подсчета для каждого субрегиона были усреднены и использовались для статистического анализа.
Влияние кетамина на более поздние JSE
Крысы, случайным образом назначенные для лечения лекарствами и стрессом, получали однократную системную инъекцию кетамина (10 или 40 мг / кг, т.е.p.) или транспортное средство за неделю до IS или HC. Через 24 часа после завершения IS или HC крысы прошли тестирование JSE.
Влияние кетамина на стресс-индуцированную активацию Fos в DRN. то же время для крыс, назначенных HC. Крысам была сделана транскардиальная перфузия 100 мл ледяного 0,9% физиологического раствора, сразу после этого — 250 мл 4% параформальдегида в растворе 0.1 М фосфатный буфер (PB; pH ~ 7,4). Мозг подвергали последующей фиксации в течение ночи в том же фиксаторе и переносили в 30% раствор сахарозы в 0,1 М PB, затем хранили при 4 ° C до разделения на срезы. Корональные срезы головного мозга, содержащие DRN, были получены при размере 35 мкм. Ткань DRN, используемую для IHC, помещали непосредственно в 24-луночный планшет.
ИГХ для Fos и 5-HT
Окрашивание для Fos и 5-HT проводили, как описано ранее (Grahn et al., 1999). Окрашивание на Fos проводили с использованием метода авидин-биотин-пероксидазы хрена (ABC).После серии промываний в 0,1 М PBS срезы инкубировали в 0,9% растворе перекиси водорода для подавления эндогенных пероксидаз. Затем срезы инкубировали в течение 24 часов при комнатной температуре с первичным антителом Fos (1:15 000; Santa Cruz Biotechnology) в блокирующем растворе, содержащем 2% нормальной козьей сыворотки (NGS), 0,5% Triton X-100 и 0,1% азида натрия. После инкубации первичного антитела срезы инкубировали в течение 2 ч при комнатной температуре в биотинилированном козьем вторичном антителе против кролика (1: 200; Jackson ImmunoResearch) в блокирующем растворе.После серии промываний PBS срезы инкубировали в ABC в течение 1 ч при комнатной температуре. Затем срезы промывали 0,1 м PB и затем подвергали воздействию раствора, содержащего 3,3-диаминобензидин (DAB), хлорид кобальта, сульфат никель-аммония, хлорид аммония и глюкозооксидазу в PB. Пероксидазную реакцию инициировали добавлением раствора глюкозы, который реагировал с тканью в течение ∼7–10 мин. Реакцию останавливали промыванием срезов PBS. Ткань помещали на предметное стекло и снимали крышку для последующего анализа.Окрашивание 5-HT проводили с использованием метода пероксидазно-антипероксидазного (PAP) метода. После серии промываний в PBS избыточное фоновое окрашивание 5-HT предотвращалось инкубацией срезов в блокирующем растворе в течение 0,5 часа. Затем ткань инкубировали в блокирующем растворе антитела 5-HT (кроличьи поликлональные 1: 10 000; Jackson ImmunoResearch) в течение 48 часов при комнатной температуре. Вторичное козье антитело против кролика (1: 200; Jackson ImmunoResearch) наносили на ткань на 2 часа после серии промывок PBS. За этим этапом последовала еще одна серия промывок PBS и инкубации с антителом PAP (1: 200; Jackson ImmunoResearch) в течение 2 часов.После серии промываний в PBS ткань инкубировали в растворе, содержащем DAB и глюкозооксидазу. Пероксидазную реакцию инициировали добавлением глюкозы и продолжали в течение 15 мин. После последней серии промывок PBS ткань помещали на предметные стекла и давали высохнуть в течение ночи. Слайды закрывали Permount.
Анализ изображений
Срезы мозга наблюдали с помощью микроскопа светлого поля (Olympus BX-61, Olympus America) и анализировали с помощью программного обеспечения cellSens (Olympus America).Все цифровые изображения были получены с использованием объектива 20x. Изображения DRN были получены с использованием параметров, аналогичных описанным ранее (Grahn et al., 1999). Срезы, соответствующие AP-координате -1,36, -1,00 и -0,70 мм относительно интераурального нуля, были взяты для рострального, среднего и каудального DRN соответственно. Fos-положительные ядра в каждой подобласти DRN наблюдались как темно-коричневые или черные круглые / яйцевидные пятна. Окрашенные 5-HT клеточные тела наблюдались в виде светло-коричневых частиц с неокрашенными ядрами и без них.Совместная локализация Fos и 5-HT наблюдалась в виде светло-коричневого клеточного тела с черным ядром. Для каждого субъекта были взяты два отдельных подсчета из разных срезов в каждой подобласти DRN. Два подсчета для каждого субрегиона были усреднены и использовались для статистического анализа.
Влияние кетамина на активацию пути PL-DRN
Микроинъекция ретроградного индикатора
Небольшое окно (1 × 1 мм) просверлили в черепе, и красные флуоресцентные RB (Lumafluor) были микроинъектированы в DRN (AP: -8.0 мм относительно брегмы, DV: -6,7 мм от поверхности черепа, ML: ± 0,0 мм относительно средней линии) с использованием шприца Hamilton на 10 мкл и металлической иглы 31-го калибра со скошенным под 45 ° кончиком. Общий объем впрыска (0,3 мкл) и скорость потока (0,1 мкл / мин) контролировали с помощью микроинжекционного насоса (UMP3-1; World Precision Instruments). Этот объем инъекции был выбран на основе пилотных экспериментов, в которых наблюдалась устойчивая экспрессия RB в пределах PL через две недели после инъекции 0,3 мкл при сохранении локализованных инъекций.После инъекции иглу оставляли на месте еще на 10 мин, чтобы обеспечить диффузию RB, после чего иглу извлекали. Небольшой разрез на черепе закрыли с помощью Vetbond (3M). Всем крысам вводили предоперационный антибиотик (Комби-Пен-48, 0,25 мл / кг, п / к) и послеоперационный анальгетик (мелоксикам, 2 мг / кг, п / к). Через две недели после микроинъекции RB крысам вводили системную инъекцию кетамина или физиологического раствора, и через 2 часа они были умерщвлены. Микроинъекции RB считались успешными, если экспрессия явно ограничивалась DRN в коронарных срезах мозга, полученных после завершения эксперимента.Для статистического анализа использовали только крыс с точными инъекциями RB.
Тканевый препарат
Крыс подвергали глубокой анестезии пентобарбиталом натрия (65 мг / кг) через 2 часа после инъекции кетамина или физиологического раствора. Крысам выполняли транскардиальную перфузию 100 мл ледяного 0,9% физиологического раствора, сразу после чего вводили 250 мл 4% параформальдегида в 0,1 М PB (pH ~ 7,4). Мозг подвергали последующей фиксации в течение ночи в том же фиксаторе и переносили в 30% раствор сахарозы в 0,1 М PB, затем хранили при 4 ° C до разделения на срезы.Корональные срезы головного мозга, содержащие PL, получали при длине волны 35 мкм. Ткань PL, использованную для IHC, помещали непосредственно в 24-луночный планшет. Ткань DRN помещали на предметные стекла и закрывали с помощью монтажной среды VectaShield (Vector Labs).
Влияние кетамина на нейронные ансамбли PL во время первоначальной инъекции и во время более позднего стресса
Вирус
Векторы аденоассоциированного вируса (AAV) были использованы для нацеливания на гибридный трансген RAM-NLS-mKate2 (RAM) нейронам. Кассеты RAM были упакованы в векторы AAV, серотипированные белками оболочки AAV1 (титры: 2.18 × 10 13 копий генома / мл) Вировека. Кассеты hSYN-eYFP (eYFP), упакованные в векторы AAV, серотипированные белками оболочки AAV1, были (титры: 3,86 × 10 12 копий генома / мл) Университетом Пенсильвании. Перед инъекцией RAM смешивали с eYFP (8,5 мкл RAM / 1,5 мкл eYFP). eYFP выражается в отсутствии нейрональной активации и служит методом определения точности инъекции после завершения эксперимента. Это гарантирует, что в отсутствие выражения RAM может быть достигнута проверка инъекции.
Доставка вирусного вектора
Крыс помещали на доксициклиновую пищу (DOX; 200 мг / кг, BioServ) за 24 часа до операции, как описано ранее (Sørensen et al., 2016). В день операции крыс анестезировали и одну одностороннюю инъекцию RAM направляли в PL (AP: +2,5 относительно брегмы, DV: -2,0 относительно пиальной поверхности, ML: ± 1,0 мм), используя параметры инъекции, аналогичные те, что описаны выше. 1000 нл ОЗУ было доставлено со скоростью 100 нл / мин. После завершения инъекционную иглу оставляли на месте еще на 10 минут, чтобы обеспечить распространение вируса, после чего иглу извлекали.Послеоперационный уход осуществлялся, как описано выше.
Мечение нейронных ансамблей во время инъекции кетамина и последующего стресса
После инъекции RAM крысы оставались на корме DOX (200 мг / кг) в течение 96 часов. Этого периода времени достаточно, чтобы предотвратить базальную индукцию RAM в отсутствие значимого события, такого как инъекция лекарства. Корм DOX был отменен и заменен стандартным лабораторным кормом через 96 часов. Удаление DOX-корма <96 ч предотвращает экспрессию RAM из-за подавления циркулирующего DOX-опосредованного трансгена (данные не показаны).Крысы оставались спокойными в колонии в течение 96 часов после отмены DOX. После 96-часового интервала DOX- крысам, рандомизированным для лечения препаратом и стрессом, вводили либо низкие дозы кетамина (10 мг / кг, внутрибрюшинно), либо высокие дозы кетамина (40 мг / кг, внутрибрюшинно), либо физиологический раствор. Через двадцать четыре часа крыс снова помещали на корм DOX (1000 мг / кг). Этот временной интервал обеспечивает устойчивое выражение RAM, сводя к минимуму неспецифическое выражение RAM. Через 48 часов крысы получали IS или HC и перфузировали через 2 часа.
Препарат ткани
Крыс подвергали глубокой анестезии и перфузии, как описано выше. Корональные срезы головного мозга, содержащие PL, получали при длине волны 35 мкм. Ткань PL, использованную для IHC, помещали непосредственно в 24-луночный планшет. Флуоресцентную ИГХ выполняли, как описано выше.
Влияние ингибирования PL-DRN на эффекты стрессовой буферизации кетамина
Вирус
Кассеты AAV2-retro-eSyn-eGF-T2A-iCre-WPRE (Cre) упаковывали в векторы AAV, серотипированные с белками оболочки AAV (титры: копий генома / мл) от Vector Biolabs.Кассеты AAV8-hSyn-DIO-hM4Di (G i ) -mCherry (hM4Di) упаковывали в векторы AAV, серотипированные белками оболочки AAV8 (титры: 4,3 × 10 12 копий генома / мл) с помощью Addgene. Кассеты AAV-hSyn-DIO-mCherry (mCherry) упаковывали в векторы AAV, серотипированные белками оболочки AAV5 (титры: 2,1 × 10 13 копий генома / мл) от Addgene.
Доставка вирусного вектора
В черепе просверлили маленькое окно (1 × 1 мм) и Cre микроинъектировали в DRN (AP: -8.0 мм относительно брегмы, DV: -6,7 мм от поверхности черепа, ML: ± 0,0 мм относительно средней линии) с использованием шприца Hamilton на 10 мкл и металлической иглы 31-го калибра со скошенным под 45 ° кончиком. Общий объем впрыска (1 мкл) и скорость потока (0,1 мкл / мин) контролировали с помощью микроинжекционного насоса (UMP3-1; World Precision Instruments). После инъекции иглу оставляли на месте еще на 10 мин, чтобы обеспечить распространение вируса, после чего иглу извлекали. Небольшой разрез на черепе закрыли с помощью Vetbond (3M).Всем крысам вводили предоперационный антибиотик (Комби-Пен-48, 0,25 мл / кг, п / к) и послеоперационный анальгетик (мелоксикам, 2 мг / кг, п / к). Через пять дней с использованием идентичных хирургических процедур hM4Di или mCherry были микроинъектированы в PL (AP: +2,5 мм относительно брегмы, DV: -2,0 мм от поверхности пиальной поверхности, ML: ± 0,5 мм относительно средней линии).
Проверка hM4Di
Чтобы определить, достаточно ли доставки CNO для предотвращения активации нейронов PL-DRN, крысы, нацеленные с помощью hM4Di на путь PL-DRN, получали однократную инъекцию CNO (3.0 мг / кг, внутрибрюшинно) или носителя за 30 минут до однократной инъекции кетамина (10 мг / кг, внутрибрюшинно). Через 90 минут крысам была сделана перфузия. Корональные срезы головного мозга, содержащие PL, получали при длине волны 35 мкм. Ткань PL, использованную для IHC, помещали непосредственно в 24-луночный планшет. Была проведена флуоресцентная ИГХ и исследована экспрессия Fos в пути PL-DRN. В статистический анализ были включены только крысы с точной двусторонней экспрессией hM4Di.
Определение эстрального цикла
Промывание влагалища проводилось перед стрессовой терапией.Пипетку с тупым концом, заполненную небольшим количеством 0,9% стерильного физиологического раствора, вводили во влагалище. Жидкость быстро вытеснялась два-три раза, чтобы осторожно смыть и собрать вагинальные клетки (~ 0,25 мл). Каплю помещали на предметное стекло и сразу же рассматривали с помощью линзы объектива 40x. Характерные изменения цитологического внешнего вида образцов использовали для определения стадии цикла: диэструс I / II (наличие ядерных клеток и лейкоцитов), проэструс (присутствие ядерных клеток) и течка (присутствие безъядерных плоскоклеточных клеток).
Молчание PL-DRN
Крыс, нацеленных с помощью hM4Di или mCherry на путь PL-DRN, случайным образом распределяли для лечения лекарствами и стресса. Крысы получали однократную системную инъекцию кетамина или носителя за неделю до IS или HC. Крысы получали однократную инъекцию CNO (3,0 мг / кг, внутрибрюшинно) за 30 мин до обработки IS или HC. Через 24 часа после завершения IS или HC поведение оценивалось во время JSE. После завершения поведенческого тестирования крыс подвергали глубокой анестезии и перфузии с использованием физиологического раствора и 4% параформальдегида.Корональные срезы, содержащие PL и DRN, помещали непосредственно на предметные стекла, и была подтверждена точная вирусная экспрессия в пути PL-DRN. В статистический анализ были включены только крысы с точной двусторонней экспрессией hM4Di или mCherry.
Статистика
Анализ данных проводился с помощью программного обеспечения Prism (GraphPad). Эффект лечения анализировали с помощью непарного теста t (лекарство) или одностороннего (стресс), двухстороннего (стресс и лекарство) или трехстороннего (стресс и лекарство и вирус) дисперсионного анализа.Основные эффекты и взаимодействия считались статистически значимыми, если p <0,05. При необходимости, апостериорных анализов и запланированные сравнения были выполнены с использованием метода Тьюки апостериорных . Значения на графиках представлены как среднее ± SEM. Результаты всех статистических анализов перечислены в статистике.
Результаты
Влияние системного кетамина на активацию JSE и DRN 5-HT
Поведение
Крысы получали либо низкие дозы кетамина (10 мг / кг, т.е.p.), высокие дозы кетамина (40 мг / кг, i.p.) или физиологический раствор (i.p.) за 7 дней до IS или HC ( n = 9–10 / группа). Тревожное поведение оценивали во время JSE через 24 часа. На рисунке 1 показано общее время, потраченное на взаимодействие в течение 3-минутного теста JSE. До введения низких или высоких доз кетамина в HC крысы не влияли на JSE, по сравнению с контрольными HC, которым вводили физиологический раствор. JSE был значительно снижен у крыс, которым ранее вводили физиологический раствор или высокие дозы кетамина, которые получали IS. Важно отметить, что низкие дозы кетамина, введенные за неделю до этого, полностью блокировали влияние IS на JSE.Двусторонний дисперсионный анализ выявил значимые основные эффекты стресса ( F (1,52) = 21,13, p <0,0001), лекарство ( F (2,52) = 12,62, p < 0,0001), и стресс от лекарственного взаимодействия ( F (2,52) = 3,97, p = 0,025). Метод Тьюки post hoc показал, что крысы, которым вводили физиологический раствор или высокие дозы кетамина перед IS, были неотличимы друг от друга, но отличались от всех других групп ( p <0.05). Крысы, которым вводили низкие дозы кетамина и IS, значительно отличались от крыс, получавших физиологический раствор и IS или высокие дозы кетамина и IS, в то время как крысы IS, которым ранее вводили низкие дозы кетамина, были неотличимы от крыс HC, которые ранее получали физиологический раствор, низкие дозы кетамина или высокие дозы кетамина. Социальное исследование было значительно снижено у крыс, которым вводили физиологический раствор и IS ( p = 0,0009) или высокие дозы кетамина и IS ( p = 0,03) по сравнению с их соответствующей группой домашней клетки.
Рисунок 1.Низкие дозы кетамина защищают самок крыс от последующего стресс-индуцированного дефицита JSE. A , Крысы получали низкую (10 мг / кг, внутрибрюшинно) или высокую (40 мг / кг, внутрибрюшинную) дозу кетамина за одну неделю до стресса или лечения HC. JSE измеряли через 24 часа после стресса или лечения HC. Метод Тьюки post hoc : * p <0,05, *** p <0,001. Столбцы представляют собой групповое среднее ± SEM.
DRN 5-HT активация
Чтобы определить, предотвращает ли кетамин типичную IS-индуцированную активацию DRN, мы оценили иммунореактивность Fos (Fos +), иммунореактивность 5-HT (5-HT +), а также процент 5-HT клеток. экспрессия Fos (% 5-HT + клеток, экспрессирующих Fos) в ростральной, средней и каудальной DRN крыс, которым за неделю до этого инъецировали низкие дозы кетамина или носителя (рис.2). Мы сконцентрировались конкретно на этих трех субрегионах DRN, потому что предыдущие исследования продемонстрировали, что у самцов крыс IS активирует среднюю и каудальную DRN (Grahn et al., 1999). Точно так же 2-часовой момент времени был выбран на основании предыдущих экспериментов, демонстрирующих индуцированное стрессом Fos в DRN (Grahn et al., 1999). На Фигуре 2 A показана типичная микрофотография, обозначающая 5-HT + клетку, Fos + клетку и 5-HT + клетку, также экспрессирующие Fos. Двусторонний дисперсионный анализ был использован для проверки того, варьировалось ли общее количество 5-HT + клеток между группами в ростральной, средней или каудальной DRN.Никаких различий в общем количестве 5-HT + клеток не наблюдалось ( p > 0,05) между группами стресса для ростральной, средней или каудальной подобластей DRN (данные не показаны). Fos + клетки исследовали в каждой подобласти DRN для различных групп обработки (фиг. 2 B , слева). В ростральной DRN двухфакторный дисперсионный анализ выявил основной эффект лекарственного средства ( F (1,38) = 4,29, p = 0,046) и основной эффект стресса ( F (1, 38) = 15.22, p = 0,0004). Апостериорный анализ Тьюки показал, что крысы IS показывают увеличение Fos + по сравнению с крысами, получавшими HC ( p <0,05). Fos + был значительно снижен у крыс, которым ранее вводили кетамин, которые получали IS, по сравнению с крысами, которым вводили физиологический раствор до IS ( p <0,05). Внутри среднего DRN стресс увеличивал Fos +; однако этот эффект был частично предотвращен предшествующим кетамином. Эти наблюдения были подтверждены двусторонним дисперсионным анализом, который выявил основные эффекты стресса ( F (1,39) = 14.94, p = 0,0004), лекарство ( F (1,39) = 4,539, p = 0,04) и взаимодействие стресс × лекарство ( F (1,39) = 4,303 , p = 0,045). Post hoc анализ продемонстрировал, что Fos + значительно увеличивался у крыс IS по сравнению с крысами HC ( p <0,01). Кетамин, введенный до IS, снижал Fos + по сравнению с крысами IS, которые получали физиологический раствор ( p <0,05). Количество Fos + -клеток в каудальной DRN было больше после IS, чем HC.Это было подтверждено двухфакторным дисперсионным анализом, который выявил основные эффекты стресса ( F (1,36) = 9,446, p = 0,004). Не удивительно, что IS вызывал стойкое увеличение количества 5-HT + клеток (5-HT + Fos) в ростральной, средней и каудальной DRN (Fig. 2 B , right). В ростральной DRN двухфакторный дисперсионный анализ выявил основные эффекты стресса ( F (1,38) = 20, p <0,0001) и лекарственного средства ( F (1,38) = 4 .255, p = 0,046). По сравнению с крысами HC, которым ранее вводили физиологический раствор, крысы IS, которым вводили физиологический раствор, показали усиление 5-HT + клеток, также экспрессирующих Fos ( p <0,05). Количество 5-HT + клеток, также экспрессирующих Fos, было снижено у крыс IS, которые ранее получали кетамин, по сравнению с крысами IS, которые получали физиологический раствор ( p <0,05). В пределах среднего DRN двухфакторный дисперсионный анализ выявил основные эффекты стресса ( F (1,39) = 26,58, p <0,0001), лекарство ( F (1,39) = 5.048, p = 0,03) и взаимодействие стресса × лекарственное средство ( F (1,39) = 5,037, p = 0,03). Как наблюдалось в ростральной DRN, крысы, которым ранее вводили физиологический раствор или кетамин, которые получали IS, показали увеличение количества 5-HT + клеток, также экспрессирующих Fos, по сравнению с крысами, которым вводили физиологический раствор или кетамин и HC ( p <0,0001). Предыдущий кетамин снижал индуцированное IS усиление 5-HT + клеток, также экспрессирующих Fos, по сравнению с крысами, обработанными физиологическим раствором, которые получали IS ( p <0.05). Наконец, в каудальной DRN двухфакторный дисперсионный анализ выявил основной эффект стресса ( F (1,36) = 23,01, p <0,0001). Крысы, обработанные физиологическим раствором, которые получали IS, показали усиление 5-HT + клеток, также экспрессирующих Fos, по сравнению с крысами HC, обработанными физиологическим раствором и кетамином ( p <0,001). Таким образом, IS активировал нейроны DRN 5-HT, и эта активация была существенно ослаблена введением кетамина 7 дней назад.
Рисунок 2.Низкие дозы кетамина подавляют активацию DRN, вызванную стрессом. A , Типичная микрофотография, показывающая 5-HT + клетку (красная стрелка), Fos + клетка (черная стрелка) и дважды меченая клетка, экспрессирующая 5-HT и Fos (белая стрелка). B , Общее количество клеток Fos + (слева) и процент 5-HT + клеток, также экспрессирующих Fos в ростральной, средней и каудальной подобластях DRN. Метод Тьюки post hoc : * p <0,05, ** p <0,01, *** p <0,001, **** p <0.0001. Столбцы представляют собой групповое среднее ± SEM.
Влияние кетамина на активность PL во время стресса
Предыдущая работа демонстрирует роль нейронов глубокого слоя (Layer V / VI) PL в буферных эффектах как кетамина, так и поведенческого контроля (Amat et al. , 2005, 2016; Варела и др., 2012). Таким образом, мы стремились определить, может ли предшествующий кетамин вызывать активацию нейронных ансамблей в пределах PL, чтобы более поздний опыт с IS теперь активировал PL в манере, совместимой с поведенческим контролем у мужчин (Baratta et al., 2009). Такие результаты позволяют предположить, что защитные эффекты кетамина опосредуются, по крайней мере частично, стойкими изменениями в нервном узле, которые обычно изучаются в отношении ингибирующего контроля над DRN (Amat et al., 2005; Baratta et al., 2009) . Рисунок 3 A показывает схематическую диаграмму экспериментальной временной шкалы. Во-первых, мы попытались определить, может ли однократная инъекция кетамина (фиг. 3 B ) вызывать экспрессию RAM (RAM +), что свидетельствует об активации PL (фиг. 3 C , D ).Чтобы оптимизировать зависимую от активности экспрессию RAM, мы оценили два разных временных интервала между удалением DOX и инъекцией кетамина. Используя этот подход, мы обнаружили, что крысы должны быть отключены от DOX (DOX-) в течение 96 часов перед инъекцией кетамина, чтобы наблюдать за маркировкой RAM. Стоит отметить, что у крыс, которым вводили DOX за 48 часов до кетамина, не наблюдалось изменений RAM по сравнению с крысами, получавшими физиологический раствор ( p = 0,65), что позволяет предположить, что DOX все еще присутствовал через 48 часов после удаления DOX. Высокие дозы кетамина, но не низкие дозы кетамина или физиологического раствора, индуцировали устойчивую экспрессию RAM + в PL (основной эффект препарата, F (2,31) = 10.71, p = 0,0003). Это не было изменено более поздним IS, как можно было ожидать, поскольку животных снова переводили на DOX через 24 часа после инъекции кетамина. Post hoc анализ показал, что крысы, которым ранее вводили высокие дозы кетамина, которые позже получали HC, демонстрировали повышенную экспрессию RAM + по сравнению с крысами, которым вводили физиологический раствор, которые позже получали HC ( p = 0,039). Точно так же крысы, которым ранее вводили высокие дозы кетамина, которые позже получали IS, показали повышенную экспрессию RAM + по сравнению с крысами, которым вводили физиологический раствор, которые позже получали ID ( p = 0.023). Затем был исследован общий Fos + после IS (фиг. 3 C , D ). Как описано ранее (Baratta et al., 2009), IS увеличивает количество клеток Fos + в PL. ANOVA выявил основные эффекты стресса ( F (1,31) = 43,69, p <0,0001). Крысы, которым ранее вводили физиологический раствор, которые получали более поздний IS, показали повышенное содержание Fos + в PL по сравнению с крысами, которым ранее вводили физиологический раствор, которые получали HC ( p = 0,002). Точно так же крысы, которым ранее вводили низкую дозу кетамина, которые получали позже IS, показали повышенный Fos +, по сравнению с крысами, которым вводили низкую дозу кетамина, которые получали HC ( p = 0.0004). Наконец, процент RAM + клеток, также экспрессирующих Fos, исследовали, чтобы определить, задействованы ли кетамин-индуцированные экспериментальные ансамбли во время более позднего IS (фиг. 3 C , D ). ANOVA выявил основные эффекты стресса ( F (1,31) = 8,55, p = 0,0064), лекарства ( F (2,31) = 16,31, p <0,0001) и стресс × взаимодействие лекарств ( F (2,31) = 5,031, p = 0.013). Поразительно, что IS вызывал значительное увеличение Fos + в нейронах RAM + PL, ранее активированных низкими, но не высокими дозами кетамина, по сравнению с крысами, которые получали HC и ранее физиологический раствор ( p <0,0001) и IS и ранее высокими дозами. доза кетамина ( p <0,001).
Рисунок 3.Кетамин-индуцированное мечение RAM транскрипционно активного нейронного ансамбля, который позже активируется неконтролируемым стрессом. A , Схема экспериментальной процедуры.Крысам вводили AAV-NLS-RAM-mKate2 (RAM). Через девять дней крысы получили однократную системную инъекцию низкой дозы кетамина (10 мг / кг, внутрибрюшинно), высокой дозы кетамина (40 мг / кг, внутрибрюшинно) или физиологического раствора. Спустя 72 часа крыс подвергали ИИ или оставляли в покое в своей домашней клетке. B , Схематическая диаграмма коронарного среза мозга крысы, демонстрирующая расположение инъекций RAM в PL. Подтверждение вирусной инъекции было подтверждено с помощью eYFP, и RAM + Fos были количественно определены в подобласти PL, обозначенной пунктирным прямоугольником. C , Репрезентативные изображения PL, показывающие клетки eYFP + (зеленые), RAM + клетки (красные), Fos + клетки (синие) и клетки RAM, экспрессирующие Fos (обозначены желтыми стрелками на крайней правой панели). Масштабная шкала: 100 мкм и применяется ко всем изображениям. D , увеличенное изображение PL, показывающее RAM (обозначено красной стрелкой), Fos (обозначено белой стрелкой) и ячейку RAM, выражающую Fos (обозначено желтой стрелкой). Масштабная линейка: 50 мкм. E , Количество меченых RAM клеток (слева), Fos + клеток (в центре) и процент двумерных RAM + клеток, которые также экспрессируют Fos (справа) в PL крыс, которые получали кетамин или физиологический раствор с последующим IS или лечение HC.Метод Тьюки post hoc : * p <0,05, ** p <0,01, *** p <0,001 для графика RAM + клеток и% RAM клеток, экспрессирующих Fos. Основной эффект двустороннего дисперсионного анализа: *** p <0,001 для графика Fos +. Столбцы представляют собой групповое среднее ± SEM.
Влияние кетамина на активность пути PL-DRN
Предыдущая работа в нашей лаборатории продемонстрировала, что путь PL-DRN критически участвует в стресс-буферных эффектах поведенческого контроля, который имеет несколько общих действий с проактивным кетамином.(Amat et al., 2005, 2014, 2016; Baratta et al., 2009). Целью настоящего эксперимента было установить, может ли предшествующий кетамин активировать путь PL-DRN способом, подобным описанному Baratta et al. (2009) для контроля над поведением. Рисунок 4 A показывает схематическую диаграмму процедуры впрыска. Экспрессию Fos исследовали в нейронах PL-DRN, меченных ретроградно (RB +) (фиг. 4 B ). Как описано ранее, инъекции RB в DRN давали экспрессию, ограниченную в основном глубокими слоями (V / VI) PL и IL, с некоторой экспрессией в дорсальной передней поясной извилине (Gabbott et al., 2005; Баратта и др., 2009; Gonçalves et al., 2009; Амат и др., 2016). Стоит отметить, что общее количество нейронов RB + PL не различается между группами ( p = 0,496; рис. 4 C , слева). Чтобы определить, активирует ли кетамин PL, мы проанализировали общее количество клеток Fos + у крыс, получавших кетамин или физиологический раствор ( n = 8 и n = 7, соответственно; рис. 4 C , в середине). Однократная инъекция кетамина увеличивала количество клеток Fos + в PL ( t (13) = 4.145, p = 0,001). Затем мы оценили, активирует ли кетамин PL-DRN путем количественной оценки процента DRN-проецирующих нейронов PL, экспрессирующих Fos (рис. 4 C , справа). Действительно, кетамин вызывает устойчивое увеличение активации пути PL-DRN ( t (13) = 3,453, p = 0,004).
Рисунок 4.Острый кетамин активирует путь PL-DRN. A , Принципиальная схема экспериментальной процедуры. Крысам вводили красные флуоресцентные RB в DRN.Две недели спустя крысы получили однократную системную инъекцию кетамина (10 мг / кг, внутрибрюшинно) или физиологического раствора. Через два часа крыс умерщвляли. Экспрессию RB и Fos оценивали в PL. B , Репрезентативные изображения PL, показывающие ячейки RB + (красный), Fos + ячейки (зеленые) и RB + Fos (обозначены белыми стрелками на крайней правой панели). Масштабная шкала: 50 мкм и применяется ко всем изображениям. C , Количество клеток, меченных RB (слева), клеток Fos + (в центре), и процентное соотношение клеток с двойной меткой, меченных RB, которые также экспрессируют Fos (справа) в PL крыс, получавших кетамин или физиологический раствор.Непарный тест t : ** p <0,01. Столбцы представляют собой групповое среднее ± SEM.
Влияние DREADD-опосредованного молчания пути PL-DRN на эффекты стресс-буферизации кетамина
Целью настоящего эксперимента было оценить, может ли хемогенетическая инактивация пути PL-DRN во время ИИ смягчить защитные эффекты предшествующего кетамина. Во-первых, была подтверждена способность hM4Di подавлять опосредованную кетамином активацию пути PL-DRN (рис.5). Фиг. 5 A изображает процедуру инъекции, используемую для нацеливания hM4Di на путь PL-DRN. Типичные изображения hM4Di, Fos + и hM4Di-положительных клеток, также экспрессирующих Fos, показаны на Фигуре 5 B . Общее количество экспрессирующих mCherry клеток в PL было определено количественно (фиг. 5 C , слева). Как было показано ранее, нейроны PL, проектирующие DRN, в основном локализованы в глубоких слоях PL (Gabbott et al., 2005; Baratta et al., 2009; Gonçalves et al., 2009). Общее количество клеток, меченных PL-DRN hM4Di, не различалось между двумя группами ( t (8) = 0.22, п. = 0,83; Рис.5 C , слева). В соответствии с результатами эксперимента по ретроградному отслеживанию PL-DRN, кетамин значительно увеличивал количество клеток Fos + в PL (рис. 5 C , в середине). По сравнению с крысами, которым вводили носитель, CNO значительно снижал вызванное кетамином увеличение PL Fos + ( t (8) = 4,337, p = 0,003). Затем исследовали Fos +, ограниченный путем PL-DRN (фиг. 5 C , справа). Крысы, которым вводили CNO, показали значительно более низкое содержание PL-DRN Fos +, индуцированного кетамином, по сравнению с крысами, которым вводили физиологический раствор ( t (8) = 4.051, п. = 0,004).
Рисунок 5.hM4Di предотвращает эффекты предшествующего кетамина на активацию пути PL-DRN. A , Принципиальная схема процедуры впрыска. Крысам вводили ретроградно транспортируемые векторы AAV, кодирующие Cre и eGFP. Пять дней спустя крысам вводили Cre-зависимые векторы AAV, кодирующие hM4Di-mCherry. Шкала: 250 мкм, применимая к обоим изображениям. B , Типичные изображения PL, показывающие экспрессию hM4Di-mCherry (слева), экспрессию Fos (в центре) и совместную локализацию hM4Di-mCherry и Fos (справа).Масштабная линейка: 50 мкм. C , Количество клеток, меченных hM4Di-mCherry (слева), клеток Fos + (в центре) и процент клеток, экспрессирующих hM4Di-mCherry, которые также экспрессируют Fos (справа) в PL крыс, которые получали кетамин перед CNO или носителем . Непарный тест t : ** p <0,01. Столбцы представляют собой групповое среднее ± SEM.
В отдельной когорте крыс, экспрессирующих PL-DRN hM4Di или mCherry, кетамин или физиологический раствор вводили за неделю до IS или HC. За 30 минут до IS или HC всем крысам вводили CNO (3.0 мг / кг, внутрибрюшинно). Таким образом, эксперимент представлял собой факторный план 2 (стресс) × 2 (лекарство) × 2 (вирус). Через 24 часа после IS или HC крысы подвергались тестированию на JSE. На рисунке 6 показано время, потраченное на взаимодействие во время теста JSE. Трехфакторный дисперсионный анализ показал основной эффект стресса ( F ( 1 , 83) = 46,98, p <0,0001), взаимодействие стресса × лекарственное средство ( F (1, 83) = 8,81, p = 0,004), взаимодействие стресса × вирус ( F (1,83) = 16.31, p = 0,0001), и взаимодействие стресса × лекарства × вируса ( F (1,83) = 9,97, p = 0,023). В группах HC ни кетамин, ни опосредованное DREADD ингибирование или их комбинация не влияли на социальное исследование ( p > 0,05). В соответствии с работой, выполненной на самцах (Amat et al., 2016) и самках крыс (Baratta et al., 2018), IS резко снизил социальное исследование у крыс, экспрессирующих mCherry, по сравнению с крысами mCherry, которые получали HC ( p <0.01). hM4Di не влиял на IS у крыс, которым ранее вводили физиологический раствор. То есть крысы, нацеленные на hM4Di, которые получали физиологический раствор и IS, напоминали крыс, экспрессирующих mCherry, которые получали IS и физиологический раствор ( p > 0,05). В соответствии с результатами, показанными на фиг. 1, предыдущее введение кетамина, но не физиологического раствора крысам, нацеленным на PL-DRN mCherry, предотвращало IS-опосредованный дефицит социального исследования у крыс, нацеленных на PL-DRN mCherry ( p <0,01). Поразительно, что опосредованное hM4Di ингибирование пути PL-DRN во время IS устраняет защитные эффекты предшествующего кетамина.Крысы, которые получали это лечение, теперь напоминали крыс mCherry, которые получали физиологический раствор до IS, или крыс hM4Di, которые получали физиологический раствор до IS ( p > 0,05).
Рисунок 6.hM4Di-опосредованное ингибирование пути PL-DRN предотвращает профилактическое действие предшествующего кетамина на JSE. Крысы, которым ранее вводили hM4Di или mCherry, нацеленные на путь PL-DRN, получали однократную инъекцию физиологического раствора или кетамина за неделю до IS или HC. JSE оценивался через 24 часа после IS или HC. Метод Тьюки post hoc : ** p <0.01. Столбцы представляют собой среднее значение группы ± стандартная ошибка среднего. Символы обозначают среднее социальное исследование крыс, получивших HC или IS во время диэструса, проэструса или течки.
Хотя это и не является предметом настоящего исследования, наблюдалась значительная вариабельность в социальных исследованиях крыс, которые получали HC или IS во время каждой фазы течки (диэструса, проэструса, течки) для разных групп лечения. Однако, учитывая небольшое количество субъектов для каждой стадии течки в каждой экспериментальной группе, невозможно узнать, влияет ли стадия течки на показатель поведенческого результата (рис.6).
Обсуждение
Настоящее исследование направлено на определение того, присутствуют ли профилактические эффекты кетамина, а также нейронные механизмы, которые опосредуют его эффекты, у самок крыс аналогично тому, что ранее было охарактеризовано у самцов крыс ( Амат и др., 2016). Таким образом, мы оценили различные поведенческие и нейрохимические конечные точки, которые были изучены в предыдущих исследованиях, проведенных с использованием самцов крыс, в то же время расширив предыдущие усилия путем оценки конкретных нейронных ансамблей и цепей, участвующих в эффектах кетамина.Результаты настоящих экспериментов предоставляют четкие доказательства того, что однократная инъекция низкой, но не высокой дозы кетамина, введенная за неделю до неконтролируемого стресса (IS), блокирует поведенческие эффекты IS на JSE, измеренные через 24 часа. Эти результаты согласуются с исследованиями нашей лаборатории (Amat et al., 2016) и другими (Brachman et al., 2016; McGowan et al., 2017), демонстрирующими профилактические эффекты кетамина у самцов крыс и мышей. Тем не менее, настоящее исследование является первым в своем роде, демонстрирующим профилактику кетамином у самок крыс, и предполагает, что профилактические эффекты кетамина на более поздние результаты стресса распространяются на оба пола.Это было особенно важно, поскольку поведенческий контроль, который имеет сходство с эффектами кетамина, не защищает самок крыс (Baratta et al., 2018), по крайней мере, при обстоятельствах, идентичных тем, которые используются у самцов (см. Ниже).
Значительный объем исследований, проведенных в основном на самцах грызунов, указывает на то, что изменения поведения, вызываемые ИИ, включая снижение уровня JSE, опосредуются активацией среднего и каудального 5-HT DRN и последующим высвобождением 5-HT в такие области мозга, как как BLA (Takase et al., 2004; Amat et al., 2008; Кристиансон и др., 2010; Dolzani et al., 2016). Эта интенсивная активация DRN необходима (Maier et al., 1993, 1994, 1995) и достаточна (Maier et al., 1995) для поведенческих изменений, связанных с IS. Таким образом, мы стремились определить, будет ли кетамин, введенный перед ИИ, смягчать активацию DRN, обычно связанную с воздействием ИИ. Действительно, однократная инъекция кетамина за неделю до ИИ значительно снизила вызванную стрессом активацию рострального и среднего DRN.Хотя это и не является фокусом настоящих экспериментов, возможно, что различия в активации субрегиона DRN между исследованиями женщин и мужчин связаны с половыми различиями в анатомии DRN. В то время как женщины, как сообщается, имеют более высокие базальные и вызванные стрессом уровни 5-HT (Domínguez et al., 2003; Mitsushima et al., 2006; Staiti et al., 2011), половые различия в анатомии DRN мало изучены. Действительно, одним из ограничений настоящих экспериментов является использование Fos в качестве единственного маркера активации DRN 5-HT.Действительно, Fos — это ранний ген, который используется для обнаружения недавно активированных нейронов (Sheng et al., 1990), но в отличие от микродиализа in vivo , где можно напрямую измерить внеклеточный 5-HT, Fos служит заместителем для DRN 5- Активация HT. Тем не менее, результаты экспериментов с двойной меткой ИГХ убедительно свидетельствуют о том, что предшествующий кетамин снижает активацию DRN 5-HT во время стресса.
Как уже отмечалось, здесь мы показали, что кетамин ослабляет нейрохимические и поведенческие последствия ИИ при введении за неделю до воздействия стрессора.На сегодняшний день никакие другие манипуляции не продемонстрировали способности вызывать эти эффекты у самок крыс. Обширный объем работ показал, что поведенческий контроль над стрессором предотвращает нейрохимические и поведенческие эффекты стрессора у самцов крыс (см. Обзор Maier et al., 2015). Удивительно, но защитные эффекты поведенческого контроля отсутствуют у самок крыс (Baratta et al., В печати). Эффекты стресс-буферизации поведенческого контроля, наблюдаемые у самцов крыс, были изучены в рамках парадигмы, в которой крысам обеспечивается контролирующая реакция (поворот колеса) на прекращение ударов хвостом (ускользающий удар хвостом; ES).Отдельная группа крыс получает идентичную серию ударов хвостом, запряженных коромыслом, однако они не могут контролировать какой-либо аспект удара хвостом (IS). Поведенческий контроль притупляет активацию DRN во время контролируемого стрессора и предотвращает вызванные стрессом поведенческие эффекты, обычно измеряемые через 24 часа. Как описано ранее, поведенческие эффекты IS являются результатом активации DRN и высвобождения 5-HT в области проекции, которые являются проксимальными медиаторами поведения, связанного со стрессором (Grahn et al., 2000; Takase et al., 2004; Amat et al., 2008; Кристиансон и др., 2010; Dolzani et al., 2016). У самцов крыс контроль за поведением активирует нейроны в глубоких слоях (V / VI) PL mPFC, которые преимущественно синапсы на ГАМКергические интернейроны в DRN (Hajós et al., 1998; Jankowski and Sesack, 2004; Gonçalves et al. , 2009). Таким образом, чистый эффект активации пути PL-DRN заключается в снижении высвобождения DRN 5-HT и блокаде поведенческих эффектов, связанных с воздействием стрессора (Amat et al., 2005). Кроме того, опыт использования ES предотвращает активацию DRN и поведенческие эффекты воздействия IS или другого стрессора, такого как социальное поражение через неделю (Amat et al., 2006; Amat et al., 2010). Удивительно, но поведенческий контроль не может активировать путь PL-DRN у женщин, и поэтому DRN-опосредованные поведенческие исходы не предотвращаются (Baratta et al., 2018). У самцов крыс защита, обеспечиваемая ЭС против воздействия более поздних неконтролируемых стрессоров, вызвана изменением состояния нейронов PL, проектирующих DRN, произведенных исходной ES, так что более поздний опыт с IS теперь активирует путь PL-DRN (Baratta и другие., 2009). Недавно было также показано, что поведенческий контроль вызывает пластичность нейронов глубокого слоя (V / VI) PL, которые анатомически расположены, чтобы предотвратить активацию DRN (Hajós et al., 1998; Varela et al., 2012; Amat et al., 2014). Более того, блокада пластичности внутри PL ингибиторами синтеза белка, APV и ингибиторами ERK перед ES предотвращает защиту ES от поведенческих эффектов более позднего IS (Amat et al., 2006; Amat et al., 2014; Christianson et al. ., 2014). Следует отметить, что эффекты кетамина в низких дозах отличаются от эффектов других антагонистов рецепторов NMDA, таких как APV (для обзора см. Abdallah et al., 2015) .⇓
Подобно эффектам контроля поведения у самцов крыс, однократная субанестетическая инъекция кетамина усиливает синаптическую пластичность в слое V PL (Li et al., 2010) и предотвращает (Li et al., 2010; Amat et al., 2016; Brachman et al., 2016; McGowan et al., 2017) и обращает вспять (Li et al., 2011) эффекты воздействия стрессора. Кроме того, кетамин, доставляемый самцам крыс, вызывает активацию PL Fos (Fuchikami et al., 2015; Amat et al., 2016) и изменяет нейроны PL-DRN, так что опыт с IS теперь активирует этот путь (Amat et al., 2016). Учитывая сходство между эффектами поведенческого контроля и кетамина на более поздние исходы ИИ, мы стремились определить, изменяет ли кетамин активность PL таким образом, чтобы более поздний опыт IS активировал ансамбль нейронов PL, первоначально активированных кетамином. Используя RAM, мы обнаружили, что кетамин в низких, но не в высоких дозах активирует ансамбли нейронов, которые позже активируются ИГ. На сегодняшний день это наиболее убедительное свидетельство того, что изменения в специфических ансамблях нейронов PL участвуют в профилактических эффектах кетамина.Другие показали, что положительный опыт активирует нейронные ансамбли в зубчатой извилине, и реактивации этих ансамблей достаточно, чтобы обратить вспять депрессивно-подобное поведение (Ramirez et al., 2015). В будущих исследованиях следует реализовать аналогичные методологии, чтобы продемонстрировать необходимость экспериментальных ансамблей в профилактических эффектах предшествующего кетамина.
Учитывая влияние кетамина на вызванную стрессом активацию DRN и активность PL во время инъекции лекарства и позже IS, мы стремились определить, активирует ли острая инъекция кетамина путь PL-DRN и активирует ли этот путь время более позднего ИС необходимо для защитного действия препарата.Результаты исследования ретроградного отслеживания показали, что острая инъекция кетамина активирует путь PL-DRN. Используя генетическую стратегию двойного пересечения вирусов, мы обнаружили, что опосредованное DREADD ингибирование пути PL-DRN во время ИИ предотвращает защитные эффекты кетамина, введенного неделей раньше. Другие выявили цепи mPFC-гиппокампа, участвующие в острых эффектах кетамина, вводимого незадолго до поведенческого тестирования (Carreno et al., 2016), однако это первое исследование, которое выявило точную нервную цепь, которая опосредует профилактические эффекты кетамина у женщин.Следует отметить, что хотя неспецифические нервные и поведенческие эффекты, связанные с метаболизмом CNO в экспериментах DREADD, были отмечены (Gomez et al., 2017), все крысы в настоящем эксперименте получали CNO, поэтому очевидно, что метаболит CNO не является посредничество наблюдаемых эффектов на поведение.
В совокупности настоящие эксперименты показывают, что кетамин оказывает длительное профилактическое действие против пагубных последствий стресса у самок крыс. Эффекты кетамина у крыс сохраняются намного дольше, чем период полувыведения из плазмы, составляющий ~ 2 часа (Williams et al., 2004), и поэтому наблюдаемые эффекты нельзя отнести к острым эффектам лекарств во время поведенческого тестирования. Структура данных предполагает, что эффекты кетамина отражают длительную профилактику, которая происходит у самцов крыс, которым обеспечен контроль над поведением. Эти данные свидетельствуют о том, что, хотя контроль поведения не может защитить самок крыс, необходимые схемы устойчивости существуют и задействованы кетамином. Следовательно, кетамин может оказаться эффективным в качестве терапевтической стратегии для женских клинических групп, склонных к высоким уровням стресса или травм.
Благодарности
Благодарности: Мы благодарим Изабеллу Фэллон и Марианн Реддан за техническую помощь и Мэтта Помренце за содержательные обсуждения и интеллектуальные комментарии.
Сноски
Авторы заявляют об отсутствии конкурирующих финансовых интересов.
Эта работа была поддержана грантами Национального института здравоохранения T32 MH016880 (для S.D.D.), R01 MH050479 (для S.F.M.) и R21 Mh206817 (для M.V.B.).
Synthesis
Рецензент Редактор: Хорхе Дж.Palop, Gladstone Institutes и UCSF
Решения обычно являются результатом того, что рецензирующий редактор и рецензенты собираются вместе и обсуждают свои рекомендации до достижения консенсуса. Когда предлагается внести изменения, ниже приводится сводное заявление, основанное на фактах, с объяснением их решения и указанием того, что необходимо для подготовки пересмотра.
Вот несколько предложений:
— Обоснуйте абстрактно клиническую значимость проведения этих экспериментов на женщинах.
— Добавьте в подписи к рисункам статистические тесты для указанных звездочек на графиках.
— Соответствуйте меткам на графиках и в тексте. Например, на рисунках нечетко используются ячейки Fos-ir, ячейки Fos + или Fos. Также будьте последовательны с метками оси Y, используются как «Fos +», так и «Fos-ir cells». Добавьте «ячейки», если «ячейки» — это единица измерения. Для поведения на Рисунке 1 используются «Домашняя клетка» и «Стресс», а на Рисунке 6 используются «HC» и «IS».
— Рисунок 1, результаты и легенда к рисунку описывают высокие (40 мг / кг) и низкие (10 мг / кг) дозы кетамина, но Рисунок 1 показывает только одну группу, получавшую кетамин.Пожалуйста, исправьте это.
— Статистика на Рисунке 1 не совпадает. На графике есть три звездочки и одна звездочка в легенде. Легенда гласит: «По сравнению с HC», но график показывает сравнение с кетамином. Измените этот рисунок, чтобы результаты, условные обозначения и графики совпадали.
— Рисунок 2, укажите дозу кетамина в результатах. Это высокая или низкая доза?
— Рисунок 3, добавьте буквы панели (A, B, C, D и E) для обозначения панелей.
— Рисунок 3C, eYFP или GFP? Пожалуйста, сопоставьте текст и легенду.Шкала указана, но не указана.
— Рисунок 3D, перед количественной оценкой опишите панель желтыми и белыми стрелками.
— Рисунок 4, добавьте буквы панели (A, B, C, D и E) для обозначения панелей. Пожалуйста, сравните статистику, **** отсутствуют в легенде.
— Рисунок 5, проверьте, составляет ли масштабная линейка 50 мкм, как указано в легенде.
(PDF) Предыдущий опыт и понимание — пример из цепи постоянного тока
Международная конференция по физическому образованию (ICPE) 2018
Journal of Physics: Conference Series 1512 (2020) 012004
IOP Publishing
doi: 10.1088 / 1742-6596 / 1512/1/012004
5
или светодиод. Его опыт работы с лампочкой в лаборатории показывает, что если ее подключить к батарее с замкнутым контуром,
, независимо от полярности батареи, она загорится. В повседневной практике ни одно устройство с батарейным питанием
не будет работать, если мы изменим полярность батареи. Этот опыт интерпретируется как «лампа
имеет полярность». Он также имел опыт работы с проводом под напряжением. Если вы коснетесь живого провода, вы получите
удара током.Эта идея привела к тому, что он ответил на вопрос, связанный с потоком заряда, что заряд
будет течь до конца резистора и рассеиваться.
Другой студент, который успешно нарисовал замкнутую цепь с помощью схемного резистора, имел только
опыта работы с лампой с байонетным цоколем, а не с лампой с винтовой цоколем. По его мнению, лампочка
не активируется, потому что соединения на схемах не к нижней части лампы. По его мнению, оба соединения
должны быть к нижней части лампы, сторона лампы должна просто удерживать лампу.
Таким образом, предыдущий опыт учащихся не помогает им понять теорию замкнутой цепи,
, скорее, они запускают разные сигналы из разных контекстов. Мы предлагаем, чтобы идея «непрерывного замкнутого контура»
преподавалась студентам в качестве основного требования для любой цепи постоянного тока для функции
в любом контексте.
Ссылки
[1] Д. Э. Браун, Дж. Клемент и У. Национальный научный фонд округа Колумбия., «Преодоление
заблуждений в механике: сравнение двух стратегий обучения на основе примеров
», Ежегодное собрание Американской ассоциации исследований в области образования,
1987.
[2] А. Карамазза, М. Макклоски и Б. Грин, «Наивная вера в« изощренных »субъектов:
неправильных представлений о траекториях объектов», т. 9, pp. 117–123, 1981.
[3] Л. К. Макдермотт, М. Л. Розенквист и Э. Х. Ван Зи, «Студенческие трудности в соединении
графиков и физики: примеры из кинематики», Am.J. Phys., Т. 55, нет. 6, pp. 503–
513, 1987.
[4] Д. Хаммер, «Ресурсы для студентов для изучения вводной физики», Am. J. Phys., Т. 68, стр.
S52, 2000.
[5] А.Э. Хаммер, «Использование эпистемологических ресурсов для изучения физики», J. Learn.
Наука, т. 12, вып. 1, 2003.
[6] Д. Хаммер, А. Элби, Р. Э. Шерр и Э. Ф. Редиш, «Ресурсы, кадрирование и передача»,
Transf. Учить. из…, с. 1–26, 2005.
[7] Д.Э. Мельцер и Р. К. Торнтон, «Информационное письмо ALIP – 1: инструкция активного обучения по физике
», Am. J. Phys., Т. 80, нет. 6, стр. 478, 2012.
[8] J. P. Smith III, A. a. ДиСесса и Дж. Рошель, «Воссоздание неправильных представлений: конструктивистский анализ знаний в переходный период», J. Learn. Sci., Т. 3, вып. 2, pp.
115–163, 1993.
[9] W. Potter et al., «Шестнадцать лет совместного обучения посредством активного осмысления
физики
(CLASP) в Калифорнийском университете в Дэвисе», Am .J. Phys., Т. 82, нет. 2, стр. 153–163, февраль 2014 г.
[10] И. Джон и С. Элли, «Цепи постоянного тока: I. Доказательства мелкозернистой контекстной зависимости »,
Eur. J. Phys., Т. 38, нет. 38, 2017.
[11] И. Джон, С. Элли, «Цепи постоянного тока: II. Определение опорных идей в цепях постоянного тока »,
Eur. J. Phys., Т. 38, нет. 38, 2017.
[12] А. ди Сесса, «К эпистемологии физики», Cogn. Инстр., Т. 10, вып.